

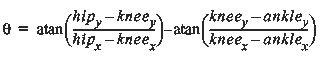
























Articles and Papers
Download this paper (1.5MB, PDF format; right-click and choose 'Save As')
Toward a Resolution of the Bigfoot Phenomenon
J. Glickman
Part 1 | Part 2 | Part 3
The Bigfoot phenomenon may be the result of a combination of sociological origin, physical manifestation through willful manufacture, and the by-product of cataloged and uncataloged animals. Observational data related to the Bigfoot phenomenon is presented and analyzed to identify its origin. Human and animal archetypes are used to demonstrate the inclusion or exclusion of these archetypes within the observations. An argument of continuity, the expectation that there may be a continuous record of the existence of an organism, is employed to include or exclude the possibility that the observations originate from an uncataloged animal. The plausibility of an uncataloged animal is examined through ecological analogy.
Monsters, and more specifically myths of Big Hairy Monsters (BHM), are a world-wide anthropological phenomenon. In North America, one such myth, centered principally in the Pacific Northwest, is known as Bigfoot. Many contemporary stories relate individual and group experiences with the Bigfoot phenomenon. Robert Pyle aptly observed, "...the phenomenon of Bigfoot exists." [Pyle 1995]. This single, lucid observation, which differentiates the existence of a Bigfoot from the existence of the phenomenon, forms the basis of this paper. Since we know that the phenomenon exists, what is its source?
The Bigfoot phenomenon may be of sociological origin, it may be physically manifested through elaborate manufacture, or it may be the by-product of an animal, cataloged or uncataloged. Its magnitude and distribution however, are, in the author's opinion, unusual and therefore important to understand. If the phenomenon is of social origin, how did it become so widespread, how does it sustain itself, and why has it been so long-lived? If the phenomenon is of elaborate manufacture, how was geographically and temporally widespread manufacture accomplished and concealed? If the phenomenon is the by-product of a cataloged animal how did human perceptual mechanisms fail? Finally, if the phenomenon is the by-product of an uncataloged animal why is there a dearth of evidence and why are we reluctant to investigate the phenomenon? Whichever of these are eventually proven to be the origin of the Bigfoot phenomenon, humanity will be the beneficiary of its investigation, by gaining new insights into the human animal.
This paper reviews observations of the phenomenon and proposes a methodology for its continued examination. A null hypothesis for this paper is formulated and presented. The observations are cataloged and their sources critiqued, which is followed by the analysis of the observations. From this analysis, new hypotheses are postulated. The conclusion presents the results of this study and provides recommendations for future studies.
Methodology
The methodology that will be used to determine the source of the Bigfoot phenomenon is:
1. Assert that there is a Bigfoot phenomenon.
2. Create a set of hypotheses enumerating the possible sources of the Bigfoot phenomenon. These include, but are not limited to, the social hypothesis, the manufacture hypothesis, the misidentification hypothesis and the uncataloged animal hypothesis.
3. Collect observations. A set of observations have been collected to facilitate the initial analysis of the phenomenon.
4. Analyze the observations to test the hypothesis.
5. Formulate new hypotheses as appropriate.
One argument that is employed to contradict the null hypothesis is the continuity argument. Continuity is an expression of evolution. Relative to the human experience, evolution is a slow process. Species gradually evolve from one to another, and eventually become extinct. There are exceptions, for example cataclysms that create adaptation challenges. Those species able to adapt survive, and those unable to adapt perish.
Some species leave a complex record of their existence, which begins with fossil evidence. Since the advent of man, extant species leave an anecdotal record through man's collective memory.
There are exceptions to both of these. For example, the chimpanzee and gorilla have no fossil record [Jones 1992] and since the beginning of this century seven new species of land mammal have been discovered [NYT 1994]. Therefore, gaps in the record of a species do not constitute unequivocal proof of non-existence.
Nonetheless, these are the exceptions and not the rule. The likelihood of a large North American animal having remained uncataloged and having no fossil record is slim. This is the essence of the continuity test: To make a plausible argument for an uncataloged animal, its continuity may be demonstrated. To demonstrate the possible implausibility of an uncataloged animal, one may illustrate discontinuities in the record.
Hypothesis
The null hypothesis has been carefully chosen because the existence of Bigfoot can not be proven due to the absence of a type specimen, therefore a null hypothesis that requires proof of the existence of Bigfoot is fatally flawed.
Archetypes do exist for proving that observations are manufactured by humans. The null hypothesis must be one that can be successfully contradicted, which may only be done with the human archetype. Thus the null hypothesis must be "The Bigfoot phenomenon originates from an uncataloged animal" because this can be contradicted by proving, for example, that an image captured on movie film is that of a human in a costume. The null hypothesis is:
* The Bigfoot phenomenon originates from an uncataloged animal.
The observations will be used to refute the null hypothesis. If the null hypothesis is successfully contradicted, then by implication:
* The Bigfoot phenomenon is of social origin,
* The Bigfoot phenomenon is the result of misidentification,
or
* The Bigfoot phenomenon is manufactured.
The Bigfoot phenomenon may originate from the super-position of observations traceable to multiple hypotheses.
Analysis
Observations of the Bigfoot phenomenon are presented, some of which are circumstantial, and among which there may be coincidence. Since there are no theories yet to model these observations, a danger resides in ascribing meaning to outcomes that are unexpected, for which an as yet absent theoretical model would predict.
Purported observation of the Bigfoot phenomenon include sightings, footprints, sounds, smells, thrown objects, hair, feces and photographs. Several individuals in the Bigfoot research community have attempted to support the phenomenon by trying to correlate the contemporary phenomenon with the European settler's historical record, Native American cultural memory, and the fossil record and are categorized as historical anecdotes.
These will be reviewed in the following sections. Things sensed (seen, heard, smelled, etc.) and subsequently reported without physical record, such as sightings, footprints, sounds, smells and thrown objects are categorized as contemporary anecdotes. In some cases, the individual or group reporting the observation presents a physical record of the event in the form of samples, footprint casts, or photographs. These materials cannot be proven to be authentic, nor do they prove the existence of an uncataloged animal because of the absence of a type specimen. These are categorized as contemporary physical observations.
Categories of observations of the Bigfoot phenomenon are shown in Table 1.
Observations from these classifications are presented in reverse temporal order — from the most recent observations to the oldest observations. Ecological plausibility and BHM as an anthropological phenomenon will be analyzed.
Contemporary Anecdotes
There are many stories, centered principally in the Pacific Northwest, that relate contemporary individual and group experiences to the Bigfoot phenomenon. Many individuals and groups comprise the Bigfoot research community, including Professor Grover Krantz, John Napier, John Green, Ray Crowe, Rene Dahinden, Bob Titmus, Ivan Sanderson and Peter Byrne to name a few. All have made some effort to collect anecdotal observations. In two cases the author is aware of, efforts have been made to formalize the collection of anecdotal observations. One such effort was led by John Green and the other by Peter Byrne.
Table 1: Categories of Observations Time (inclusive) Category Examples
Contemporary
(postdate 1958) Anecdotes sightings, sounds, footprints, smells, thrown objects
Physical Record footprint casts, hair samples, photography (film, video, still)
Historical
(predate 1958) Anecdotes settler historical record, Native American cultural memory
Physical Record fossils
The role of the contemporary anecdotal observations is to support or refute the main hypothesis. Each qualified anecdote is quantified by representing the anecdote as a geo-time coded event, i.e. date, time, latitude, longitude and altitude. This dataset is then analyzed by SPSS 1, a computer-based statistical analysis software package.
Green's Sighting Data
John Green has been involved in the Bigfoot community for approximately thirty years and as of the 1981 printing of his book [Green 1981] claimed to have over 1,500 confirmed sightings. Mr. Green's current data was not formally made available to this study, so the methods employed by him and the manner by which his data are organized cannot be assessed.
As an alternative to using his current data, Green's national sighting data as of November 1977 is summarized in Table 2 [Green 1981]. Green's data is analyzed first because it covers the largest geographic area, and to the best of the author's knowledge, is the only collection of continental data.
Methodology
Green's data will be tested against a simplistic model of expected sighting rates for animals. The probability of receiving a report for a cataloged animal is modeled as:
Pr = Ps. Pa . Ph . Pe (Eq.1)
where,
Pr is the probability function of receiving a report,
Ps is the probability function that an observation results in a report submission,
Pa is the probability function of an animal being at a specific place and time to be observed,
Ph is the probability function of a human being in a specific place and time to make the observation, and
Pe is the probability function of an observer expecting to observe the phenomenon.
The author assumes that the probability that an observation results in a report submission is geographically uniform, so this reduces to a constant. The probability of an animal being in a specific place and time to be observed is directly proportional to the animal's population density. A uniform distribution is assumed. In the event the animal's population density is non-uniform, this becomes superimposed upon the result. The probability that a human in a specific place and time makes an observation is directly proportional to human population density. This is modeled on a per-state basis as the number of square miles divided by the population [Gousha 1995].
Analysis
Table 2 is organized on a per-state basis and is ordered in descending normalized frequency. The "Freq." column contains Green's reported observation frequencies [Green 1981]. "Dist." is an ordinal distance reference as measured from the geographic center of the state to the geographic center of Washington. "Sq. Mi." is the number of square miles in the state. "Population" is the 1980 population census figure for the state. "Pop./Sq. Mi." is derived as "Population" divided by "Sq.Mi." "Norm. Freq." is the normalized frequency and is derived as "Freq." divided by "Pop./Sq.Mi."
Therefore:
Equation 2Eq. 2
"Group" is the assigned cluster group resulting from cluster analysis (presented below). Canadian data is not included, due to incomplete data.
Table 2: Green Sighting Data Case State Dist. Freq. Sq.Mi. Human
Population Pop/Sq.
Mi. Norm
Freq. Cluster
Group
1 Alaska 76 20 550,000 400,481 0.73 27.47 A
2 Montana 22 74 147,138 786,690 5.35 13.84 A
3 Oregon 10 176 96,981 2,632,663 27.15 6.48 A
4 Washington 0 281 68,192 4,130,163 60.57 4.64 A
5 N.California(Est.) 25 294 79,347 5,917,141 74.57 3.94 A
6 S.California(Est.) 35 49 79,347 17,751,422 223.72 0.22 B
7 Idaho 15 32 83,557 943,935 11.30 2.83 A
8 Wyoming 31 4 94,914 470,816 4.96 0.81 B
9 South Dakota 44 7 77,047 690,178 8.96 0.78 B
10 Nevada 26 5 110,540 799,184 7.23 0.69 B
11 New Mexico 52 7 121,510 1,299,968 10.70 0.65 B
12 Florida 107 104 58,560 9,739,992 166.33 0.63 B
13 Texas 70 30 267,339 14,228,283 53.22 0.56 B
14 Arkansas 74 19 53,104 2,285,513 43.04 0.44 B
15 Iowa 60 15 56,290 2,913,387 51.76 0.29 B
16 North Dakota 40 2 70,665 652,695 9.24 0.22 B
17 Arizona 45 5 113,575 2,717,866 23.93 0.21 B
18 Kansas 55 6 82,264 2,363,208 28.73 0.21 B
19 Oklahoma 64 9 69,919 3,025,261 43.27 0.21 B
20 Mississippi 83 8 47,716 2,520,638 52.83 0.15 B
21 Nebraska 48 3 77,227 1,570,006 20.33 0.15 B
22 Colorado 42 4 104,247 2,888,834 27.71 0.14 B
23 Missouri 67 10 69,686 4,917,444 70.57 0.14 B
24 Maine 105 4 33,040 1,124,660 34.04 0.12 B
25 Utah 32 2 84,916 1,461,037 17.21 0.12 B
26 Illinois 71 23 56,400 11,418,461 202.45 0.11 B
27 Michigan 75 18 58,216 9,258,344 159.03 0.11 B
28 Georgia 95 10 58,876 5,464,265 92.81 0.11 B
29 Minnesota 53 5 84,068 4,077,148 48.50 0.10 B
30 Indiana 77 15 36,291 5,490,179 151.28 0.10 B
31 Wisconsin 64 8 56,154 4,705,355 83.79 0.10 B
32 Pennsylvania 93 24 45,333 11,866,728 261.77 0.09 B
33 Tennessee 84 9 42,244 4,590,750 108.67 0.08 B
34 Kentucky 84 7 40,395 3,661,433 90.64 0.08 B
35 West Virginia 90 6 24,181 1,949,644 80.63 0.07 B
36 Ohio 84 19 41,222 10,797,419 261.93 0.07 B
37 Alabama 88 5 51,069 3,890,061 76.17 0.07 B
38 South Carolina 98 6 31,055 3,119,208 100.44 0.06 B
39 Louisiana 82 5 48,523 4,203,972 86.64 0.06 B
40 New Hampshire 102 5 9,304 920,610 98.95 0.05 B
41 North Carolina 99 5 52,712 5,874,429 111.44 0.04 B
42 New Jersey 101 36 7,836 7,364,158 939.79 0.04 B
43 Vermont 99 2 9,609 511,456 53.23 0.04 B
44 New York 95 11 49,576 17,557,288 354.15 0.03 B
45 Virginia 96 4 40,815 5,346,279 130.99 0.03 B
46 Maryland 98 12 10,577 4,216,446 398.64 0.03 B
47 Delaware 100 1 2,057 592,225 287.91 0.00 B
48 Connecticut 103 2 5,009 3,107,576 620.40 0.00 B
49 Massachusetts 102 1 8,257 5,737,037 694.81 0.00 B
50 Rhode Island 105 0 1,214 947,154 780.19 0.00 B
Mean 69.32 28.18 71,362 4,497,982 147.05 1.35
Median 75.50 7.50 56,345 3,113,392 75.37 0.12
Std. Dev. 4.18 61.09 11,613 601,667 206.58 4.39
Std. Err. 29.53 8.64 82,114 4,254,426 29.22 0.62
Table 3 presents bivariate correlation coefficients for Table 2 data between frequency and population, and frequency and population density are computed as a baseline prior to data clustering and is called the baseline correlation.
The frequency is not well correlated to either the population or the population density across the entire dataset. Hierarchical cluster analysis was subsequently performed on the normalized frequency. Clustering was done by case, and a range of solutions from two to five clusters was computed. The result of cluster analysis is presented in Table 4.
The lack of additional cases in cluster group Green5 from cluster group Green4 suggests two things: that the cases in clusters 1 through 4 of cluster group Green5 are differentiated from the rest of the dataset, and that two clusters is the appropriate cluster size since the hierarchical analysis simply rearranged the set of cases in Green4 and Green5.
Cases 1, 2, 3, 4, 5 and 7 are called Group A which consists of Alaska, Montana, Oregon, Washington, Northern California and Idaho. The remainder of the cases are called Group B. The "Cluster Group" column in Table 2 shows the result of clustering.
The same correlations as those computed for the baseline were computed for Group A and B and are summarized in Table 5.
Discussion
The relationship in the clustered data is the correlation between population density and frequency: the Group A correlation of +0.9661 is high relative to the Group B correlation of +0.1244.
A second relationship in the clustered data is the correlation between population and frequency. When Group A is separated from the dataset, its correlation to population rises from +0.1192 to +0.5664.
Group A is differentiated from Group B by its high correlation to population density. This is consistent with the model of receiving a report of a cataloged animal (Eq. 1).
Table 3: Correlation of Green's Data to Population Statistics Frequency vs. Population Frequency vs. Population Density
Baseline Correlation +0.1192 +0.2673
Significance 0.410 0.061
Cases 50 50
Table 4: Cluster Analysis of Green Sighting Data Cluster Group
Name Number of
Clusters Cluster1 Cluster2 Cluster3 Cluster4 Cluster5
Green2 2 1 all others N/A N/A N/A
Green3 3 1 2 all others N/A N/A
Green4 4 1 2 3,4,5,7 all others N/A
Green5 5 1 2 3 4,5,7 all others
Let's assume that manufactured reports will be uniformly distributed across the population. If the rate of manufactured reports is constant, then the frequency of reports should correlate to population. To some degree, this is seen in Group B. There may be other unidentified influencing factors such as mean media exposure to Bigfoot, which may influence the density of manufacturing. The author speculates that Group A and Group B represent different phenomenon. Group B may represent manufactured reports because of the correlation to population, whereas Group A may represent a different phenomenon because of its correlation to population density. The author hypothesizes that if Green's data is the superposition of multiple phenomena that this is the expected result.
Sapunov reports a theory of testimonies developed and employed in the USSR in the mid 1980s capable of testing populations of eyewitness reports for authenticity:
The mathematical theory of testimonies was developed mainly on data from traffic incidents (Rossinsky 1984). According to the theory, the distribution of quantitative characters of observed items within a group of witnesses must be normal or Guassian. Subjective biases on the part of witnesses tend to displace the mode of distribution. The qualifications or educational backgrounds of witnesses influence the variance of distribution: the higher the qualifications or education, the less is the variance of distribution. [Sapunov 1988]
Sapunov continues:
According to the theory of testimonies, the extremes of the quantitative traits reported by a group of independent witnesses should be distributed in the tail or tails of a normal or Guassian distribution if the data are authentic (Rossinsky 1984). False reports would be distributed with many peaks, and without tails. The existence of one or two modes suggests a single direction of hoaxing — which is unlikely — or the objective reality of the reports. [Sapunov 1988]
Table 5: Post-Clustering Correlations of Green's Sighting Data to Population Statistics Frequency vs. Population Frequency vs. Population Density
Baseline Correlation +0.1192 +0.2673
Baseline Significance 0.410 0.061
Baseline Cases 50 50
Group A Correlation +0.9626 +0.9661
Group A Significance 0.002 0.002
Group A Cases 6 6
Group B Correlation +0.5664 +0.1244
Group B Significance 0.000 0.421
Group B Cases 44 44
TBRP GIS† 1 Data
(†Geographic Information System)
Peter Byrne has been in the Bigfoot community on a full-time basis for seventeen of the last thirty-five years most recently serving as the Director of The Bigfoot Research Project (TBRP). Whereas Green's data is national with coarse geographic information, TBRP's data is regional with precise geographic information. Based on Byrne's intuition, TBRP focused solely on the Pacific Northwest. In so doing, TBRP was investigating the Group A phenomenon. While this permitted TBRP to study that region in more depth, it is also unfortunate that there is no national data with which to compare to their regional results.
TBRP collected ancedotal observations by soliciting reports via a toll-free telephone number through newspaper advertisements. During the month of May 1996, TBRP received two-thousand-two-hundred-sixty telephone calls, most of which were categorized as nuisance calls from children. Since 1992, TBRP has collected approximately one-thousand regional anecdotal observations, three-hundred and seventy-four of which have been deemed credible by TBRP, though the methodology by which this determination was made is subjective.
Methodology
When TBRP received a non-nuisance telephone call it identified what type of anecdotal observation was being reported and filled out a survey form specific to this type. There was one survey used for sightings (15 pages), one for footprints (11 pages), and a combined survey for sounds, smells and thrown objects (12 pages). The surveys were authored by TBRP and were not examined by a survey professional for bias or leading questions.
A subset of these anecdotes were geocoded and entered into a computer database (This dataset is referred to as TBRP1 and is shown in Figure 1). TBRP staff employed an informal model of what constituted a credible report which they developed intuitively. The credibility of an anecdote was assessed by the subjective application of this informal model. If the anecdote matched their informal model closely enough, it was deemed credible. This method filtered the anecdotes according to TBRP staff expectations and skewed the computer database toward the staff's informal model. Anecdotes were further categorized with a credibility rating of "A" through "C" based upon the personal judgment of TBRP staff.
A limited amount of information was entered into a computer database, which included a case number, date of occurrence, location description, latitude, longitude, altitude, one or more anecdote classifications consisting of sighting, footprint, sound, smell, or thrown object, and the credibility rating. As of June 17th, 1996, three-hundred and seventy-four anecdotes were cataloged by TBRP as credible, all of them in the Pacific Northwest. One-hundred and sixty-seven of these have complete information including date, altitude, and geocoding. These one-hundred and sixty-seven reports, which are referred to as Group I, are the dataset for the analysis below.
Definitions of Anecdotal Classifications
There are five anecdotal classifications recognized by TBRP. These are sightings, footprints, sounds, smells and thrown objects. Anecdotes are cataloged as a:
* Sighting when the observer reports seeing a Bigfoot. If a photograph is presented the case is still given a sighting classification.
* Footprint when the observer reports seeing a large footprint. If a plaster cast of one or more of the footprints is presented the case is still given a footprint classification.
* Sound when the observer reports loud whistling, screaming, or roaring.
* Smell when the observer reports an overpowering, noxious smell.
* Thrown Object when the observer reports objects thrown.
Whenever more than one classification is applicable, multiple classifications are associated with the case.
Fig. 1
Figure 1: GIS Data
Fig. 2
Figure 2: GIS Analysis
Analysis
TBRP's geocoded data was analyzed for patterns. Correlation coefficients were computed for all pairs of latitude, longitude, altitude, month and year in the dataset that had complete information. No significant correlations were found.
A new dataset was created, containing twelve cases, one for each month (This dataset is referred to as TBRP2). Frequency data by month, mean monthly latitude, mean monthly longitude and mean monthly altitude were aggregated from dataset TBRP1 and entered into dataset TBRP2. Mean monthly temperature and mean monthly precipitation for Portland, Oregon were manually added to dataset TBRP2. Correlation coefficients were computed for all pairs in dataset TBRP2. The only significant correlations found were between mean latitude, mean longitude and mean altitude, suggesting that there is a geographic pattern to the location of the reports filed with TBRP. This geographic pattern could be correlated with where the population lives, where people misidentify animals, where people are seeing an uncataloged animal, etc.
Figure 2 shows a high density of reports in and near Hood River County, Oregon. While the hot spot toward the center appears to be reporting the bias, the diagonal band from the upper right to the lower left is of interest. This area corresponds to the maximum altitude portion of the Cascade range to the south and west of Cascade Locks, Oregon, and to the north and east of Stevenson, Washington and Carson, Washington. These areas are very rugged and inaccessible. It is interesting to note that this high density area of reports originates from a low-population density area.
Fig. 3
Figure 3: Scatter Diagram of Latitudes and Longitudes Articles and Papers
Download this paper (1.5MB, PDF format; right-click and choose 'Save As')
Toward a Resolution of the Bigfoot Phenomenon
J. Glickman
Part 1 | Part 2 | Part 3
Contemporary Physical Record
The purported physical record of the Bigfoot phenomenon consists of footprint casts, photographs, hair and feces samples.
The most extensive collection of footprint casts that the author is aware of belongs to Dr. Grover Krantz, a Professor of Anthropology at Washington State University. As of 1992 Professor Krantz reports having eighty-one plaster casts which he believes represent twenty-two separate individuals [Krantz 1992]. Professor Krantz also reports that John Green and Bob Titmus have the casts of approximately another twenty individuals. These collections were not examined by the author during the course of this research.
Dr. Henner Fahrenbach has performed statistical analyses on the footprint subset of Mr. Green's data, which to the best of the author's knowledge is the largest footprint study to date. While the author has seen Dr. Fahrenbach's results, the author has not studied Dr. Fahrenbach's methodology. Because Dr. Fahrenbach's work is based on Mr. Green's data which was not made available to this study, the author has not incorporated Dr. Fahrenbach's results [Fahrenbach 1995].
The only alternate source of large footprints that the author is aware of is the double foot registration made by bears. Novices sometimes interpret these bear tracks as "Bigfoot" footprints, although experienced trackers can readily tell them apart.
On several occasions individuals have presented hair samples for analysis. To date, and without exception, these have been determined to be synthetic fibers, the hair of previously cataloged animals, or plant matter. Several samples recently presented are currently undergoing analysis.
On several occasions individuals have presented photographic imagery for analysis, most of which, with the exception of the 1967 Patterson-Gimlin film [Patterson and Gimlin 1967], have been demonstrated as forgeries.
Image processing is applied to the Patterson-Gimlin film to search for evidence of forgery, the result of which follows. The role of the contemporary physical record in this research and more specifically the Patterson-Gimlin film, is to refute the null hypothesis, that the Bigfoot phenomenon originates from an uncataloged animal. Objective data extracted from the film will be presented, along with morphological information which is compared to the great apes.
Analysis of 1967 Patterson-Gimlin Film
The 1967 Patterson-Gimlin Film, photographed on October 20, 1967 in Bluff Creek, California by Roger Patterson and Bob Gimlin is one of few pieces of photographic imagery that has not yet been proven to be a forgery. This film was evaluated by the author to see if it may be excluded from the set of Bigfoot phenomenon observations by demonstrating it is a forgery.
Methodology
While it is not possible to prove the imagery in the Patterson-Gimlin film represents an uncataloged animal, it is possible to extract physical and biomechanical parameters which may exclude the possibility that it is a human in a costume. Analysis of the film was performed by converting it to digital data via a film scanner so that the imagery could be processed by computer. The original of the Patterson-Gimlin film is believed to have been photographed on Kodak Kodachrome film. The camera used was a Kodak K-100 16mm movie camera. A first generation copy of the film was made on Eastman 78 Safety film on November 8, 1968, a little more than one year after the original was shot. This first generation copy was first examined by the author in October, 1993 for shrinkage and brittleness to ensure it could withstand digitization. The film had been in storage for twenty-five years in the controlled environment of a bank safety-deposit box and was in excellent condition. The data in Table 6 was extracted from the film (as of October, 1993).
In May, 1994 the first generation copy was hand-carried via a commercial airline flight to Oxberry ATI, a company located in Carlstadt, New Jersey. Oxberry is a manufacturer of high-resolution film scanners that are used by the motion picture industry. One scanner Oxberry manufactures is a modular high-resolution scanner which, when the proper parts are installed, is capable of digitizing 16mm movie film. As of May, 1994 Oxberry had never sold the 16mm components, so Oxberry was contracted to assemble a 16mm scanner from inventory and digitize the film.
Each of the 953 frames of the Patterson-Gimlin film were digitized three times, once each through red, green and blue filters. Each of these three colors was digitized using 12 bits (4,096 levels) at the rate of 2,656 pixels horizontally and 1,912 pixels vertically. This provided 36 bits per pixel at a digitization rate of slightly higher than 250 dots per millimeter (approximately 6,350 dots per inch) which was high enough to image the film grain. Each frame of the Patterson-Gimlin Film yielded a 30,470,144 byte Silicon Graphics (SGI) file composed of a 512 byte header followed by the pixels organized in 3 planes, one for each color. These frame data files were buffered locally on a Silicon Graphics workstation until twenty frames were accumulated which were then written to an 8mm Exabyte tape cartridge in Unix tar format. The twenty frame files were written twice to each tape to minimize the chance of a tape defect rendering a frame inaccessible.
Table 6: Film Data Film Width(mm) Image Width(mm) Image Height(mm) Sprockets, Center-to-Center(mm)
Specification N/A 10.49 7.605 7.605
Measured 15.75 10.29 7.60 7.568
Deviation N/A -2.0% -0.07% -0.5%
Because the total size of the dataset is nearly 30 billion bytes (GB) of data, it was impractical in 1994 to keep all of the frames on rotating magnetic storage (hard disk). Instead, each group of twenty frames was written to a recordable CD-ROM. The full dataset is comprised of 48 CD-ROMs which provide near-line, random access to the dataset. All data transfers were performed with 100% read-after-write verification. The software used to perform the image restoration, image enhancement, and other image processing is Clarity, a proprietary image processing software package developed by Photek. Clarity was run on a specially built ALR Evolution V, IBM-compatible computer. Other software used includes Adobe Photoshop and Kodak Shoebox.
Most of the images presented were processed with either image restoration or image enhancement software. Some of the enhancement algorithms operate only on gray-scale, or result in gray-scale images after processing. The original scanned images are RGB, and gray-scale images are the Y component of the YIQ transform. The image contained on the first generation copy that was scanned resulted from the optical superposition of a Kodachrome original onto Eastman Safety stock. This is important because the dyes used in the film for the three colors are not the same size. For this reason, in some instances, the green layer of the film is processed alone because it has the smallest grain and hence captured the highest spatially-resolute image. The image restorations involved motion and focal blur removal which was performed using FIR and IIR filters. Image enhancement included Wallis enhancements, homomorphic equalization, histogram equalization and curve adjustments.
Analysis
Analyses can be performed on anthropometric and bio-mechanical properties of the subject observed in the 1967 Patterson-Gimlin film. Three types of analyses follow:
* Mensuration & Estimation,
* Kinematics, and
* Morphology.
All of these analyses require accurate spatial and temporal references, neither of which have been available to date. An accurate temporal reference has not been successfully established as a result of this research.
An accurate spatial reference has been established from research photographs taken by Byrne and Hodgson in 1972 at the Bluff Creek site [Byrne 1972]. While Patterson believed he set the Kodak K-100 16mm motion camera to 16 frames per second (fps) prior to his departure for Bluff Creek, he reported that in the circumstances surrounding the filming, notably the behavior of his horse, that he may have bumped the film speed selection dial. Subsequent to the filming, Patterson did not check or record the position of the film speed selection dial. Even if Patterson did know the setting, the Kodak Archives reports that the speed tolerance of the K-100's film carrier subassembly was ±10% resulting in a range of film speeds from 14.4 fps to 17.6 fps when the film speed selection dial is set to 16 fps [Kodak 1995]. An accurate spatial reference has been established from research photography taken by Byrne and Hodgson in 1972 at the Bluff Creek site. During the course of filming, Patterson filmed several dead trees some of which also appear in Byrne's 1972 research photography of the site, which is not normally useful by itself. However, also contained in Byrne's research photography is an assistant, Michael Hodgson, who is holding a vertical scale (apparently marked every six inches).
Fig. 5: Hodgson Reference
Figure 5: Hodgson Reference
Fig. 4: Hodgson Reference
Figure 4: Hodgson Reference
Hodgson reported his 1972 height as 6' 0" without shoes and weight as 150 lbs [Hodgson 1995]. Figures 4 and 5 were used to validate Hodgson's height. While Byrne took several photographs of Hodgson that day, the one shown is Figure 4 was used because Hodgson's feet were visible and Hodgson was in the same z-axis plane as the vertical scale. Even though the base of the vertical scale is not visible, the bottom of the scale was artificially extended to verify that it was in the same plane as Hodgson's feet. Because Hodgson and the vertical scale are in the same z-axis plane, Hodgson's height may be read from the scale. The photograph was digitally enhanced to make the washed-out scale markings visible, and shows Hodgson's height as 6' 1¾". Louise Robbins reports mean subject footwear height as 2", so Hodgson's height with footwear is within the expected range [Robbins 1985]. Therefore, Hodgson's total height including footwear is established as 6' 1¾".
Mensuration & Estimation
The mensuration method, which is lengthy, is not presented here for sake of brevity. Contact the author for complete information.
Height Analysis
Fig. 6: Height Analysis
Figure 6: Height Analysis
In a separate reference photograph taken by Byrne, Hodgson is seen moving through a z-axis image plane that the subject moves through in frame 326. Since the height of Hodgson has been established and because he is in the same z-axis plane as the subject, Hodgson's height may be used to measure the subject's height provided that the scale of the two photographs can be matched. Both images contain a dead tree, the size of which did not change during the intervening 5 years which is used to match the scales of the two photographs. During scaling, three points were used to validate that the tree had not changed size. Figure 6 is a digital composite of the film subject and Hodgson in the same image plane with a common image scale established by the dead tree.
Note the successful planar alignment of the feet in this matched-scale composite. Hodgson's height in pixels is 276, and the height of the film subject is 327. A simple ratio is used to compute the height of the film subject:
Equation 3 (Eq. 3)
Therefore,
Equation 4 (Eq. 4)
Thus, the computed height of the subject in the Patterson-Gimlin film is 7' 3½". An error analysis has not yet been undertaken, but in other similar forensic situations is typically ±1".
Waist Perimeter
The waist of the subject in the film is modeled as an ellipse, with the height of the film subject used as the scale reference.
The major axis of the waist ellipse was extracted from frame 61 and is 30.8". The minor axis of the waist ellipse was extracted from frame 339 and is 20.4" (Refer to [Glickman 1997] for the extraction method]). Oblique and scale corrections were required. The perimeter of an ellipse is approximated by [Hudson 1917] (Note: a is ½ of the major axis, and b is ½ of the minor axis):
Equation 5 (Eq. 5)
Equation 6 (Eq. 6)
Thus, the computed waist perimeter of the subject in the Patterson-Gimlin film is 81.3". An error analysis has not yet been undertaken.
Chest Perimeter
The determination of chest size plays an important role in estimating mass and caloric requirements. The chest of the subject in the film is modeled as an ellipse (see Waist Perimeter above), with the height of the film subject used as the scale reference.
The major axis of the chest ellipse was extracted from frame 61 and is 31.4". The minor axis of the chest ellipse was extracted from frame 339 and is 20.9" [Glickman 1997]. Oblique and scale corrections were required. The perimeter of an ellipse is defined in Equation 5. Thus, using Equation 5 the computed chest perimeter of the subject in the Patterson-Gimlin film is 83". An error analysis has not yet been undertaken.
The methodology used for the extraction of the chest perimeter is discussed at length in [Glickman 1997].
Estimated Mass
The mass of all primates has been shown to be allometrically related to chest size [McMahon 1983]. Whether this equation is applicable to the subject of the Patterson-Gimlin film is open to debate. It is nonetheless interesting to note the mass estimated by this equation.
The allometric relationship that relates chest circumference in centimeters to mass in kilograms is:
Equation 7 (Eq. 7)
Substituting the chest circumference in centimeters, the estimated mass of the subject in the Patterson-Gimlin film is 887 kg or 1,957 lbs. An error analysis has not yet been undertaken.
Stride Length
The average stride length reported by Patterson, Gimlin and Titmus measured from the footprint impressions was 81.5" [Krantz 1992]. Byrne reports that this measurement may be in error, as it may have been measured from heel to toe, or toe to heel, rather than heel to heel, or toe to toe. Frame 308 shows a half-stride of the subject.
Using the previously computed subject height as a scale reference, the half-stride length is computed as 46.7" after oblique and scale corrections. Multiplied by 2 to make it equivalent to the average stride measurement reported is 93.4" which is substantially different than that reported. Two possible explanations for this are that the footprints measured by Patterson, Gimlin and Titmus were not made by the film subject, or that an error was made by Patterson, Gimlin and Titmus in measuring and reporting the stride length. With regard to the latter, the difference between the field and film measurements is 11.9" which is close to the 14.5" length of the plaster cast footprint impressions, the difference being 2.6" or
Given the error progagation of oblique and scale corrections, this is in within the expected range. These results suggest that Patterson, Gimlin and Titmus inadvertently measured from toe to heel. An error analysis has not yet been performed.
Arm Length
The length of the subject's arm is computed from frame 326. Using the previously computed subject height as a scale reference, the subject's arm length of 161 pixels is computed as 43". An error analysis has not yet been performed.
Leg Length
The length of the subject's leg is computed from frame 326. Using the previously computed subject height as a scale reference, the subject's leg length of 150 pixels is computed as 40" accounting for the bent knee. An error analysis has not yet been performed.
The leg length expected for this height in a human is 46.4" [Winter 1990]. The standard leg to height ratio is .53H [Winter 1990]. The observed ratio extracted from the film is .46H. The leg length of the subject is 3.0 standard deviations from the human mean which is the 99.9 percentile and is present in one out of 1,000 people. While the length of the leg could be the result of a prosthesis, it is the opinion of the author that the probability of this is low because foot flexion is observed in the film.
Edge Detection
Edge detection algorithms were run on key frames, including frame 352, with the hope that if the subject was a person wearing a costume, that a seam or interface in the costume would be detected [Gonzalez 1987]. No seams or interfaces were detected.
Morphology
The morphology, or appearance of the film subject is compared to the great apes. Special attention is given to comparisons with the mountain gorilla because of the visual similarity. Hand, foot, face and body morphology are analyzed in the following sections.
Hand Morphology
The hand of the subject is seen clearly in only a few frames. Figure 7 shows the fingers extended whereas
Figures 8 and 9 show the hand clenched, thus hand flexion is evident in the film, demonstrating that the hand is not a solid, inflexible prosthesis. In Figure 10 a photograph of a clenched gorilla hand is shown for visual reference [Godwin 1994].
Fig. 7: Subject Hand
Figure 7: Subject Hand
Fig. 9: Subject Hand
Figure 9: Subject Hand
Fig. 8: Subject Hand and Fingers
Figure 8: Subject Hand and Fingers
Fig. 10: Gorilla Fingers, Detail
Figure 10: Gorilla Fingers, Detail
A series of hands from the great apes is shown in Figure 11 which illustrates the evolutionary changes from the gibbon to the human [Kogod 1993]. The notable adaptations include the shortening of the hand and fingers and the movement of the thumb upward.
Fig. 11: Hands of the Great Apes
Figure 11: Hands of the Great Apes
It is the author's opinion that the subject's hand appears to be between that of the gorilla and the human.
The arm length expected for this height in a human is 38.5" [Winters 1990]. The standard arm to height ratio is .44H [Winters 1990] (See Appendix D). The ratio extracted from the film is .49H. The arm length of the subject is 5.5 standard deviations from the human mean which is the 99.9999981 percentile or is present in one out of 52.5 million people [Weimer 1993].
This suggests that if the subject is a human in a costume that some form of arm prosthesis is in use. Finger and hand flexion is observed in the film which implies that the prosthesis must support flexion. The use of such a sophisticated prosthesis appears to be at odds with the year the film was made, the technology available at that time, and the financial resources of those involved with the filming.
Foot Morphology
Several frames of the Patterson-Gimlin film clearly image the foot. Figure 12 shows the foot above the ground, whereas Figure 13 shows the foot undergoing flexion, which demonstrates that the foot in the film is not a solid, inflexible prosthesis.
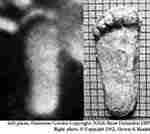
Note that separate toes are visible in the subject. Note the wide heel, minimal instep, and other key features of the foot including the position of the instep and ball below the big toe which closely resembles the plaster cast of the foot imprint taken by Titmus (Figure 14) [Krantz 1992]. This result links the plaster casts to the film subject for the first time, suggesting that the plaster cast could have been made contemporaneously by the film subject.
Fig. 12: Subject Foot
Figure 12: Subject Foot
Fig. 13: Subject Foot Flexion
Figure 13: Subject Foot Flexion
Fig. 14: Subject Foot Flexion
Figure 14: Subject Foot Flexion
A series of feet from the great apes is shown in Figure 15 which illustrates the evolutionary changes from the gibbon to the human [Kogod 1993]. The notable adaptations include the shortening of the foot and toes and movement of the big toe upward as its grasping role decreases.
Fig. 15: Feet of the Great Apes
Figure 15: Feet of the Great Apes
It is the author's opinion that the subject's foot appears to be between that of the gorilla and the human.
Face Morphology
Fig. 17: Gorilla Head, Profile Figure 17: Gorilla Head, Profile
Fig. 16: Subject Head, Profile Figure 16: Subject Head, Profile
Subjectively, the face and head appear to be gorilla-like in profile, but human-like when viewed from the front. Figure 16 shows a frontal brow, sagittal crest and an ear location that are similar in appearance to that of the mountain gorilla (Figure 17). The bulbous region around the mouth (mouth plate), is less pronounced in the mountain gorilla, and absent in the human. The visible texture on the face of the subject suggests less facial hair than the mountain gorilla, but more than a human. The jaw of the subject is below the shoulder line as in a gorilla, whereas the human jaw is above the shoulder line. When the subject is seen in motion, the shoulders are rotated when the subject turns its the head toward the camera. This is mechanically required as a result of the jaw being below the shoulder line, as in the mountain gorilla.
Fig. 18: Subject Face Figure 18: Subject Face
Fig. 19: Subject Face, Detail Figure 19: Subject Face, Detail
Figures 18 and 19 show a front-view of the subject's face which bears some subjective resemblance to a human.
It is the author's opinion that the relationship between the frontal brow, nose and mouth of the subject is between that of the mountain gorilla and the human. It is also the author's opinion that the relationship of the nose to the frontal brow-mouth plate plane places the subject between the mountain gorilla and the human.
Body Morphology
Fig. 21: Gorilla Neck and Back
Figure 21: Gorilla Neck and Back
Fig. 20: Subject Neck and Back
Figure 20: Subject Neck and Back
There are several other morphological similarities between the film subject and the mountain gorilla. In Figures 20 and 21 the configuration and appearance of the lower neck and lower back musculature of the subject is shown to be similar to the mountain gorilla. In Figures 22 and 23 the configuration and appearance of the side-torso musculature is also shown to be similar to the mountain gorilla.
There are several ways the subject may be differentiated from traditional forgeries: non-uniform hair texture, non-uniform coloration and non-uniformity of hair length. Examples of sophisticated costumes include those fabricated for the motion pictures 2001 A Space Odyssey, Gorillas in the Mist, Congo, the Planet of the Apes series as well as Eugene O'Neil's stage drama "The Great Apes". Peter Elliott, its lead, is shown in costume in Figure 24.
Fig. 22: Gorilla Torso, Side
Figure 22: Gorilla Torso, Side
Fig. 23: Subject Torso, Side
Figure 23: Subject Torso, Side
Fig. 24: Gorilla Costume
Figure 24: Gorilla Costume
Fig. 25: Gorilla and Subject breasts
Figure 25: Gorilla and Subject breasts
Most costumes have uniform hair length and little variation in color. Less sophisticated costumes have baggy, shapeless bodies with little or no detectable musculature, non-visible buttocks, and inflexible hands and feet. The appearance and sophistication of musculature as seen in the Patterson-Gimlin film has not yet been reproduced in costumes in the entertainment industry.
Figure 25 shows the breasts of a gorilla and the subject.
Kinematics
The kinematics section investigates the motion of the subject's knee as compared to that of a human and illustrates one of the muscle groups seen in motion in the Patterson-Gimlin film.
Knee Kinematics - Knee Delta
People who have viewed the Patterson-Gimlin film have commented on whether the subject's walk looks like a human walk or not. The opinions have run the gamut from statements to the effect that "it walks exactly like a human", to "it walks nothing like a human".
Fig. 26: Joint Data Extraction
Figure 26: Joint Data Extraction
These opinions have been rendered from a mixed set of film replay speeds, which is methodologically faulty because the recording speed is not known. Thus, these opinions have been purely subjective. Joint range-of-motion between the film subject and a human can be compared. While the elbow and wrist are two potential candidates, the knee was chosen because it may be the joint most differentiated from human motion parameters.
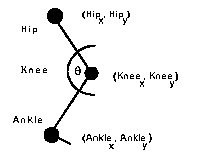
Preliminary objective gait data has been extracted from the film by digitizing the presumed positions of the hip joint, knee joint and ankle. First, the camera motion introduced by the cameraman was removed by extracting a 640 x 400 pixel subset of the 2,656 x 1,912 original relative to the hip of the subject.
The joints were digitized on-line on the computer using "play" an interactive forensic frame editing program (Figure 26) [Photek 1996].
These positions of the joints were estimated by observing the relationship of the surface deformation. For example, the hip joint was estimated by looking at the relationship of the torso to the upper leg and selecting the position that closely approximately the intersection of the medial axes. This type of estimation can be inaccurate and result in noisy data. At least four types of error can contribute to the noise including the:
* surface deformation which can cause the same surface point to change relationship with the underlying joint,
* repeatability of the surface plasticity which can change with the relationship of the underlying joint,
* subjective judgment of the human performing the digitization, who may not make repeatable judgments,
and
* perspective error resulting from the oblique angle of the subject with relationship to the camera.
Fig. 27: Joint Segments
Figure 27: Joint Segments
The data extracted was not corrected for this oblique angle so as not to introduce additional noise into the data. If the digitized joint positions were corrected for this, the recorded angles would increase. The interior angle of the knee was computed as the difference between the angles described by the hip-knee and the knee-ankle line segments (Figure 27):
Equation 8 (Eq. 8)
The knee theta for a human subject is shown in Figure 29 [Winters 1990]. The graphed data for the subject knee theta is a shown in Figure 30.
Fig. 29: Human knee theta
Fig. 30: Subject knee theta
A cycle for the human knee contains two distinct phases, a swing phase and a weight transfer phase. The knee theta of the film subject shows a more gradual transfer of weight rather than a separate phase. Assuming the subject knee delta is not overwhelmed with noise, the film subject is not employing typical human locomotion.
What is important is whether a human can replicate the knee delta observed in the film. Could a 7' 3½" tall human maintain the stable head position (relative to the ground) with the continuous forward motion observed in the film while emulating a stride length of 93". Preliminary experiments by the author suggest that while the stride can be duplicated, the continuous forward motion seen in the film can not. The knee delta data extraction must be repeated to verify the extracted graph, and the proposed experiment should be performed.
Moving Muscle Groups
In several places in the Patterson-Gimlin film, groups of muscles in motion can be seen, in the arms, back and legs. One example shown in Figure 28 is the equivalent of the quadricep muscle in the human which is seen expanding while it absorbs the weight of the subject. Also seen in this sequence is a structure similar to a knee cap, the shape of which changes like a human knee. This is particularly difficult to forge because of the need for surface conforming material. Surface plasticity in the side torso is seen near frame 352. This requires not only a conforming material, but a material with independent x and y plasticity to avoid detectable material folds.
Fig. 28: Subject Leg Muscle
Figure 28: Subject Leg Muscle
Discussion
To date the Patterson-Gimlin film has defied explanation, and it continues to do so after three years of rigorous forensic examination. If the Patterson-Gimlin film is a forgery it is the opinion of the author that it required substantial capital, which probably was not available to Patterson and Gimlin, and sophisticated knowledge of primate anatomy and materials science.
If the Patterson-Gimlin film is of an uncataloged animal, its subject bears a strong resemblance to the mountain gorilla. Most of the comparative morphological analysis place the film subject either similar to the mountain gorilla or between a gorilla and a human.
The knee kinematics of the film subject do not appear to be normal for a human, but might possibly be simulated by a 7' 3½" human. It may be possible to conclusively demonstrate that a human cannot reproduce this motion with proper experimental design. It is the author's opinion that a scientist whose discipline is biomechanics might be able to prove that the subject in the film is or is not human from the kinematics data, and in particular the knee delta.
Motions visible in the film that are difficult to replicate in a costume include the knee cap, the upper leg muscle motion, and the surface plasticity. Also present is non-uniform hair length, non-uniform hair color, and well-defined body shape.
If only a single dimension of similarity was seen in the Patterson-Gimlin film it could easily be dismissed as a forgery. In the opinion of the author, the Patterson-Gimlin film is remarkable in the simultaneous presence of all of the dimensions listed above.
Despite three years of rigorous examination by the author, the Patterson-Gimlin film can not be demonstrated to be a forgery at this time.
Part 1 | Part 2 | Part 3
Download this paper (1.5MB, PDF format; right-click and choose 'Save As
Articles and Papers
Download this paper (1.5MB, PDF format; right-click and choose 'Save As')
Toward a Resolution of the Bigfoot Phenomenon
J. Glickman
Part 1 | Part 2 | Part 3
Historical Anecdotes
European settler records and Native American mythology comprise the historical anecdotes. Newspapers of fifty to one-hundred and fifty years ago contain accounts of what today are purported to be Bigfoot observations. There are also the personal journals of trading company employees who explored and settled what is today western Canada and the United States that also contain accounts of purported Bigfoot observations. These accounts pre-date the contemporary name "Bigfoot" which entered use in 1958 [Green 1981]. These anecdotes will not be reviewed in this paper and the reader is directed to John Green's book, Sasquatch, The Apes Among Us for an informal survey of these accounts.
Several members of the Bigfoot research community, including Henry Franzoni and Gayle Highpine, have attempted to relate the contemporary Bigfoot phenomenon to Native American mythology [Franzoni 1996]. It is challenging to accommodate these inquiries in a scientific investigation for several reasons. The first is the understanding gap between European culture and Native American culture. For example, the use of the label "mythology" is really a misnomer. Native American culture is based upon verbal rather than written communication. These verbal communications are used for many purposes including what European culture calls parables, stories, myths, spirituality and explanations. We simply do not have a word in English that defines the relationship of these verbal communications to the Native American culture. For lack of a better word, the author will call them "stories." The major hurdle for the European culture in understanding Native American culture is the concept of "truth." The European culture seeks truth through scientific explanation, whereas Native American stories are truth in the Native American culture. In the context of European culture, Native American stories are mixtures of what we call reality and mythology and are used by Native Americans to explain and define the world. This makes the examination of the Native American stories for information concerning Bigfoot especially difficult, particularly because of the natural human tendency to create Big Hairy Monster (BHM) stories. This mixture of myth and reality makes it challenging to use objective western methods to evaluate story content.
Native American Stories
Henry Franzoni has surveyed Native American mythology in search of a link to the Bigfoot phenomenon [Franzoni 1996]. Most cultures have BHM myths and the Native American culture is not an exception. Because of the mixture of mythology and reality in Native American stories, the author found that most of these myths, with varying degrees of ambiguity, overlap with BHM myths. The author could not determine a way to quantify Franzoni's survey, however in an effort to follow through on Franzoni's idea, the author located a dictionary of Native American myths [Gill 1992].
This dictionary, the result of a national compilation of Native American myths, defines each myth and also lists the tribe and region of origin. Because of the large number of myths in the book, the author elected to perform a frequency analysis, by region, of Native American BHM myths contained in the dictionary.
BHM is not the only "universal" myth — for instance dwarf myths can be found in most cultures of the world as well. As a "control," the author performed a frequency analysis of Native American dwarf myths. Table 7 summarizes the frequency by region, of dwarf and giant myths in the Dictionary of Native American Mythology.
This analysis is subjective, and there are many criticisms that may be made of it. For one, the author "judged" whether a particular myth was a dwarf or a giant myth and did so without a formal methodology. Undoubtedly, someone repeating this experiment might omit a particular myth or include one that the author did not. Nonetheless, the frequency of dwarf myths is relatively evenly distributed by region. The frequency of giant myths is highest in the Northwest and Arctic regions, and with the exception of the Northeast, closely approximates the frequency of the dwarf myth. Whereas the frequency of dwarf myths may be interpreted as a basal rate, the giant myth frequency bears a resemblance to the distribution of Green's sightings data.
The higher Northwest density may be an expression of a more deeply ingrained regional cultural myth. Why then does Green's data parallel this? Are there documented cases of a myth crossing from one culture to another in a few decades? If this myth crossed from the Native Americans to the European settlers, are there others that did as well?
Henry Franzoni located a Native American, Gayle Highpine, who was also studying the relationship between the Bigfoot phenomenon and Native American mythology. She observed regional differentiation in Native American mythology — Northwest myths having physical and tangible content, whereas regions distant from the Northwest coast having more spiritual and supernatural content. The following is a quote of Highpine's writing taken from Franzoni's paper:
...I have never heard anyone from a Northwestern tribe suggest that Bigfoot is anything other than a physical being, living in the same physical dimensions as humans and other animals. He eats, he sleeps, he poops, he cares for his family members. However, among many Indians elsewhere in North America... as widely separated at[sic] the Hopi, the Sioux, the Iroquois, and the Northern Athabascan — Bigfoot is seen more as a sort of supernatural or spirit being, whose appearance to humans is always meant to convey some kind of message. [Franzoni 1996]
Table 7: Frequency of Native American Giant and Dwarf Myths Cultural Region Dwarfs Giants
Northwest 2 7
Subarctic 2 5
East Arctic 0 4
Northeast 2 4
Plains 2 3
Southeast 1 2
Great Basin 0 1
Arctic 1 1
Southwest 0 0
California 0 0
Plateau 0 0
The meaning of Native American myths can change in translation. Nonetheless, it is useful to illustrate Highpine's point with two myths, one from the Northeast and one from the Northwest.
In the Northeast the Iroquois and Onandaga tribes have a myth about a being called Dehotgohsgayeh:
This giant lives in the south along the margin of the earth in total darkness. One side of his body is red, the other black...
As translated, there are several aspects of this myth that deviate from our western understanding of the world. One can read meaning into the myth, but this is rather like fortune telling due to the ambiguity of the description — many meanings can be found depending upon what one is seeking. This then, is a good example of the difficulty of analyzing Native American myths.
In the Pacific Northwest the Coos tribe has a myth about a being called Geldegwests:
Benign giants who live near streams and eat fish.
This translation of this myth is a declarative, with what seems little "mythological" content.
Franzoni located Oregon Geographic Names in which its author, Mr. Lewis A. MacArthur, observed a faint geographic pattern between Native American and European legends [Franzoni 1996]:
Devils Lake Fork takes it's name from the fact that it drains a small body of water called Devils Lake, so called as a result of Indian nomenclature. The Indians, particularly of the Coast Range region, were fearful of a number of lakes and localities that were supposed to be inhabited by skookums, or evil wood-spirits. Some of the lakes are still called Skookum lakes, others are called Devils lakes...
Franzoni used MacArthur's hypothesis as a spring-board to develop one of his own, that contemporary geographic names might be a record of the Native American memory of Bigfoot. To pursue this inquiry, Franzoni employed the Geographic Names Information System [US Department of the Interior 1994] computer software. Franzoni developed the following search term consisting of English, Spanish and Native American names related to the Bigfoot phenomenon:
Skookum, Devil, Spirit#holy, Wampus, Splin-tercat, Windago, diablo, Sasquatch, Bigfoot, big+foot, Ape, Monkey, Wild Woman, Hock?m*, Cultus
Specific geographic features were excluded. Franzoni used the following exclusion term:
#church+#school+#airport+#building+#cemetery
698 geographic locations matched the search parameters. Table 8 presents the frequency of geographic locations grouped by state in descending order.
Table 8: GNIS Search Results State Frequency State Frequency State Frequency
Oregon 89 South Dakota 12 New York 5
Washington 70 Georgia 11 Ohio 5
California 62 Maine 11 Virginia 5
Idaho 41 Michigan 11 Massachusetts 4
Arizona 39 Missouri 11 Pennsylvania 4
Alaska 29 Kentucky 10 Wyoming 4
New Mexico 27 Arizona 9 Puerto Rico 4
Minnesota 26 Florida 9 Indiana 3
Nevada 23 Iowa 7 Maryland 3
Utah 22 Louisiana 7 Kansas 2
Wisconsin 22 Tennessee 7 Oklahoma 2
Montana 21 West Virginia 7 Wash. DC 1
Texas 21 Alabama 6 Hawaii 1
Colorado 17 South Carolina 6 Illinois 1
N. Carolina 14 Mississippi 5 New Jersey 1
Discussion
The GNIS search results parallel the Native American mythology distribution and Green's sightings data.
As suggested by MacArthur, the geographic place names identified by the GNIS search may be a reflection of Native American mythology, and therefore the parallel to Native American myth distribution is expected. While the parallel may be intriguing, it can be independent of the Bigfoot phenomenon and therefore does not contribute to identifying whether the phenomenon is of social or physical origin.
Understanding the relationship between the GNIS results and Green's data, is more difficult. Most of Green's data is from after 1958, when the American public was slowly becoming aware of the Bigfoot phenomenon, which suggests there is probably little influence from Native American culture. Because the original claims of sightings were in the Pacific Northwest, expectations may have been set that sightings should occur in the Northwest skewing the geographic distribution of "sighting" reports to the present day. Thus, the parallels seen between the GNIS results and Green's data may be coincidence.
Certainly the European settlers had no expectation of encountering the Bigfoot phenomenon, yet within the first few decades of their North American arrival reports originate from the first trade company representatives. During the late 1700s and the 1800s there are similar reports, principally west of the Rockies. It is important for us to remember that Bigfoot was not generally known back then as there was no large scale media for distribution as we have today, however, we must keep in mind that BHM as an anthropological phenomenon should have been as common then as it is now. The reports from the 1700s and 1800s have only recently been found, after 1958 when Bigfoot became popularized in North America.
Historical Physical Record
The lack of a type specimen and a fossil record may defeat the continuity argument. The proper conclusion is that the phenomenon does not originate from an uncataloged animal.
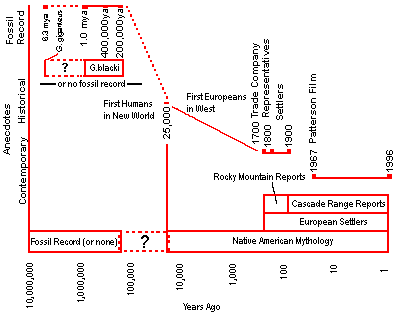
Several individuals have speculated that the existing fossil record of Gigantopithecus may be the related to the Bigfoot phenomenon — that perhaps an uncataloged descendant of Gigantopithecus is the source of the Bigfoot phenomenon.
Such an assertion is highly speculative, the plausibility of which is examined by reviewing the fossil evidence of Gigantopithecus and its presumed ecology and relating them to the anecdotal observations.
Paleontology
The continuity argument is derived from the expectation that there may be a continuous record of an organisms existence: fossils during human pre-history, written and verbal records for as long as humans have had language, and for extant species, live specimens.
Of the existing fossil record, Gigantopithecus is the only candidate fossil that approximates the size of the reported anecdotal observations. Gigantopithecus is known only by four mandibles and approximately one-thousand teeth — no post-cranial material has been found. Three of the four mandibles, known as Gigantopithecus blacki, are approximately 300,000 to 400,000 years and are from the Kwangsi Province of southern China. The fourth mandible, known as Gigantopithecus giganteus, is from the Siwalik Hills of Bilaspur, north of Delhi in India and is approximately 6.3 million years old. It is smaller than the three Chinese mandibles and is believed to be an older species of the same genus. The Gigantopithecus teeth were discovered in Liucheng, Kwangsi, Wuming, Bama, Daxin, and Jianshi of southern China. Because of the lack of post-cranial material, the posture and locomotion of Gigantopithecus is unknown.
Anthropologists believe that Gigantopithecus became extinct in the middle Pleistocene, at the latest between 200,000 and 400,000 years ago.
The taxonomy of Gigantopithecus has been controversial — originally thought to be hominid, then pongid, later ramapithecene. Recently, the consensus that Ramapithecus is a female Sivapithecus suggests that Gigantopithecus is related to pongo via a common ancestor, Sivapithecus. The contemporaneous discovery of pongo fossils at Gigantopithecus sites may support this. Today, the majority of anthropologists believe Gigantopithecus was pongid and not hominid:
If synapomorphies are correctly identified, the orangutan is the sole living descendant of the once successful Sivapithecus group, which was not ancestral to later African hominoids. The enigmatic Chinese Pleistocene hominoid Gigantopithecus blacki may be similarly derived... [Delson 1985]
Discussion
Some species leave behind records in the form of fossils, although few individual animals are converted to fossils. There are several possible reasons why fossils of Bigfoot have not been found:
* Non-existence. The phenomenon does not originate from an uncataloged animal.
* Environment. Certain environments are more likely to support fossil formation than others.
* Misclassification. Existing fossils attributed to an inappropriate genus or species.
* Undiscovered. Fossils exist but have not been unearthed.
The process of fossilization does not convert all deceased animals to fossils — most decompose before they can be fossilized because specific environmental conditions are required to create a fossil. Fortey explains fossil formation:
All fossils are found in rocks that were originally unconsolidated sediments... Certain environments which today support a rich and varied plant and animal life have no sediments forming in them, and the organisms living there have virtually no chance of being preserved in the fossil record. Mountainous regions, for example, are dominated by the erosion of the rock forming the ranges, and therefore no permanent sediment is formed there. Torrential rain and rapid weathering, aided in some climates by the action of frost, rapidly destroys much of the organic material: the chances of any preservable remains reaching a lowland river where permanent sediment is accumulating are remote. The faunas and floras of mountainous regions of the past are most unlikely to be represented in the fossil record. The fossilization potential of a mountainous environment is low. [Fortey 1991]
Thus, where the deposition of undisturbed sediment dominates, fossils may form. Where erosion dominates, such as the montane, fossils rarely form. Suspending disbelief momentarily, of the sightings deemed credible by TBRP, most are in the montane environment. Asian reports, such as the so-called Yeti of the Himalayas, are from a similar environment. If these are sightings of an uncataloged animal, then such an environment would rarely produce a fossil.
When the environment of an animal is restricted to a sufficiently small region, and if this region does not support fossil formation, a gap in the fossil record of an animal may form.
The fossil record of ape evolution is confined almost entirely to the Miocene epoch, from 23 million to 5 million years ago... Ape lineages did persist into the Plio-Pleistocene, although some subsequently became extinct. All these surviving lineages were probably more widespread than they were today. However, their record after about 8 million years ago includes only scanty remains of a recently extinct giant ape (Gigantopithecus) and Pliocene fossils of uncertain affinity, all from southeastern Asia. There is no fossil record of chimpanzees or gorillas at all. [Jones 1992]
Science accepts the existence of the gorilla and chimpanzee through the observation of type specimens even though there is no fossil record. As a single dimension, the lack of fossil evidence does not constitute conclusive proof of an animal's non-existence.
Paleontologists believe Gigantopithecus became extinct between 400,000 and 200,000 years ago, in part because this is the age of the youngest Gigantopithecus fossil. Gigantopithecus however has a scant fossil record that contains a gap of approximately 5 million years between the Indian mandible and the oldest Chinese tooth. Such gaps are typical of the fossil record. Even the intensely investigated hominid record has a 3 million year gap [Tattersall 1993]. Given the 5 million year gap in the Gigantopithecus fossil record, it seems presumptive to interpret a 400,000 year absence of fossils as conclusive proof of the species being extinct. Nonetheless, it is proper scientific procedure to do so until new hard evidence becomes available.
Misclassification of a fossil may occur through the premature introduction of a new taxonomic class, by the introduction of a taxonomic class too late, or through the misattribution of a fossil of one taxonomic class to another. An example is the 1915 discovery of a lower molar by Pilgrim (GSI D-175) that was attributed to Dryopithecus giganteus. Von Koenigswald discovered a large tooth in 1935 in an apothecary shop in China and suggested the genus Gigantopithecus. It was only later that GSI D-175 was properly attributed to Gigantopithecus.
Paleoclimatology
Animals adapt to ecological change which is driven, in part, by changes in climate.
During the Pleistocene, when climates oscillated over many thousands of years between warm and cold, marine and land organisms migrated backwards and forwards with the climatic shifts to keep living in the conditions to which they were adapted. [Fortey 1991]
If synapomorphies are correctly identified, the orangutan is the sole living descendant of the once successful Sivapithecus group, which was not ancestral to later African hominoids. The enigmatic Chinese Pleistocene hominoid Gigantopithecus blacki may be similarly derived... [Delson 1985]
Climate is defined by the superposition of cycles, some of which may have astrophysical origins. During the 1930s Milankovitch hypothesized that fluctuations in climate are driven by variations in the earth's orbit and rotational axis. Dansgaard at the University of Copenhagen has deduced mean temperature changes by analyzing the proportion of heavy oxygen in ice core samples with a mass spectrometer. His methods, and others, have demonstrated that some aspects of Milankovitch's hypothesis, that orbital perturbations affect climate, may be true [Turekian 1971].
During the last 160,000 years, or since roughly the most recent time Ciochon believes that Gigantopithecus became extinct, the climate consisted of:
Table 9: Climate Time (Years Ago) Climate (Description)
160,000 Deep Ice Age
140,000 Rapid warming over 10,000 years to an even warmer temperature than it is today
120,000 Cooling into an ice age...
100,000 ... fluctuating ...
80,000 ... but gradually ...
60,000 ... deepening ...
40,000 ... over 100,000 years
20,000 Rapid warming over 5,000 to 10,000 years, warming to present temperatures
Discussion
Several have postulated that Gigantopithecus could not have made the crossing from Eurasia to the New World across the Bering Land Bridge because such a crossing was too arduous for a species unless it possessed a social structure and culture to facilitate cooperation. There is, however, evidence that other mammals made this crossing, presumably without such cooperation:
Additional material for the determination of climatic changes during the Late Cenozoic is provided by the intercontinental migrations of mammals between Eurasia and America (Repenning 1967). At least four periods of intense exchange between faunas of the Old and New World can be distinguished in the interval between the present day and the middle Pliocene (Hemphillian in the American stratigraphy). Without going into the paleontological details one can say that the first wave of migration indicates the occurrence of a moist and warm forest environment in the Bering Land Bridge region. The migration wave corresponding to the Villafranchian (or the Blancan in the New World) suggests the presence of a forest vegetation on the route, but with open areas and a temperate climate. The deterioration of the climate on the migration route of the mammals continued, and the great wave of migration in the late Pleistocene included only arctic species, inhabitants of steppes, tundra and, at the most, the northern zone of taiga. As time lapsed, the faunal exchange was more and more limited to one direction only. The peak was reached in the late Pleistocene; in this period 23 mammalian species passed from Eurasia to North America and none migrated from the opposite direction. [Turekian 1971]
Turekian's inference that the Bering Land Bridge region of this period was a forest environment contradicts the expectation that a period generally colder than today could support a forest rather than tundra. His inference suggests this environment was at one time similar to the environment of present day anecdotal observations. South of the Bering Land Bridge, approximately 18,000 years ago during the last ice age, a north-south corridor between the Laurentide ice sheet and the ice on the western mountains ran from what is today northern Alaska via the Canadian Rockies into the Cascades and US Rockies [Calder 1974]. This is consistent with North American sighting distributions of which there were some reports from the Rocky Mountains during the 1700s and 1800s. Nearly all current sightings in TBRP's database originate in the Cascades.
There are theories that Neanderthals were physically adapted for the colder conditions that existed at the beginning of the last glaciation. As the last glaciation ended, Neanderthals may have, as did most organisms, migrate with the climatic shift. As Homo erectus became successful in the warming climate, Neanderthals may have been assimilated into the population through interbreeding. There is some evidence to support this in the form of transitional fossils between Neanderthal and Homo erectus that are approximately 300,000 years in age. Gigantopithecus was also affected by the warming climate and may have migrated to higher altitudes or latitudes seeking the cooler conditions to which it was previously adapted.
While there have been several intervening climatic shifts, it is, nonetheless, warmer today than it was 300,000 years ago. If, as a result of this warming, Gigantopithecus migrated to a cooler and perhaps montane environment, there is little expectation of recent fossil formation.
Paleoecology
Understanding the presumed ecology of Gigantopithecus can contribute to demonstrating the plausibility or implausibility that it is related to the Bigfoot phenomenon. There is differing and conflicting opinions and evidence. Paleoecological evidence indicates that a similar ecology existed in India 6.3 million years ago and in China 400,000 to 300,000 years ago, the age of G. giganteous and G. blacki fossils:
The habitat of the Indian specimen of Gigantopithecus (named Gigantopithecus bilaspurensis), which flourished at least four million and possibly more than eight million years earlier, seems to have been somewhat like the habitat in Kwangsi. The fauna found in the early Dhok Pathan sediments... suggests that the environment consisted largely of dry grasslands. Thus both the Indian and the Chinese faunas lend support to the hypothesis that the giant apes had adapted to an open environment quite unlike the forest habitat of their fruit-and-leaf eating pongid ancestors. [Simons]
Fossilized plant remains embedded in fossilized tooth enamel lends clues as to the composition of the Gigantopithecus diet. Albeit derived from a small sample, electron microscopy has found the presence of grass and fruit phytoliths during the examination of Gigantopithecus teeth. Ciochon speculates that the grass phytoliths are from bamboo. The fruit phytoliths are believed to be from the Moraceae or a closely related family which includes the sugar maple, fig, jackfruit and durian. Since the Moraceae do not grow in the savanna or its fringe, this habitat is ruled out as the habitat of Gigantopithecus [Piperno 1990].
The evidence now emerging is that Gigantopithecus was an eclectic feeder that concentrated on fruits as well as tough, fibrous vegetation. As White (19) has aptly noted, Gigantopithecus "seems to be adapted to a diet both high in carbos or starches, and requiring heavy mastification (p. 231). [Ciochon 1990]
The dentition and jaw of Gigantopithecus suggest powerful chewing [Yinyun 1982]. Some have hypothesized that powerful chewing was an adaptation for specialized diets such as gravinominous feeding, however there is no proof of this.
Jones summarizes his opinion of the ecology of Gigantopithecus:
The diet and habitats of late Miocene Eurasian hominoids were probably quite diverse. Sivapithecus, Ouranopithecus and Gigantopithecus had thick-enamelled cheek teeth, and may have had diets similar to that of Kenyapithecus, with hard and tough food items... By the end of the Miocene, all these Eurasian lineages were extinct in the areas from which they are known as fossils, except China. Their extinction might have been caused by a cooler, drier and more seasonal world climate, which emerged as the Miocene progressed, and by the resulting decline in evergreen forest and woodland and its partial replacement with deciduous forest, scrub and, perhaps, the first extensive wooded grasslands... The lineages leading to gibbons and the orang-utan persisted, but were confined to the forests of Southeast Asia. The enormous and enigmatic Gigantopithecus was probably a ground-dweller in more open habitats before its extinction in the later Pleistocene. [Jones 1992]
Discussion
Both orangutan (pongo) and giant panda (Ailuropoda) fossils have been found contemporaneously at Gigantopithecus sites which suggests a possible similarity in ecologies among these species.
Fossil Pongo is the only higher primate usually discovered with Gigantopithecus blacki. That does not mean the ecological environments for fossil Pongo and Gigantopithecus are the same, but at least they may not be considerably different. [Yinyun 1982]
Thus, it has been suggested that Gigantopithecus might have a diet similar to the giant panda [White 1975]. Piperno's identification of phytoliths has provided the first hard evidence as to the diet of Gigantopithecus, consisting of at least grasses and fruits.
The giant panda, presumed extinct until their discovery by the west in the 1920s, have migrated from the ecology of Gigantopithecus to the montane environment.
Today the giant panda is confined to the uplands of central China, in montane forests where dense stands of Bamboo grow. [Ciochon 1990]
The giant panda's typical altitude range is 5,000 to 7,000 feet although they have been seen at altitudes up to 16,000 feet. Orangutans have adapted to varied ecologies such as the lowland and swamp forests and the montane environments of Borneo and Sumatra. The giant panda has a specialized diet with more than 99% of their food consisting of bamboo when in their natural habitat:
The koala and panda are often cited as examples of extreme food specialization. Indeed, the koala's physiology appears so adapted to a diet of Eucalyptus leaves that the animal cannot change readily to another food supply (Eberhard 1978). Pandas have obviously specialized on bamboo in various ways, but they remain essentially omnivores; their dependence on bamboo reflects mainly the lack of a large alternative food supply, especially in winter, rather than the inability to assimilate other foods. [Schaller 1985]
Pandas are now known to eat meat as well as more than 25 species of plant. When presented with meat, many pandas will immediately eat it [Schaller 1985]. This is not too surprising, considering the panda is believed to have evolved from carnivorous ancestors. The fact that they eat little meat in their natural habitat may be the result of a lack of opportunity rather than one of preference. When faced with a shortage of bamboo, pandas will eat other foods. When in captivity, pandas readily adapt to a diet of porridge [Schaller 1985]. Because of the high-bulk and low nutritional content of bamboo, and thus the need to consume large quantities of it, the giant panda spends more than 40 percent of the day at rest and is never seen running.
An outstanding characteristic of giant herbivores is their extreme slowness. They have no particular need of speed; their size and thick skins protect them from predators, and of course their feeding habits require no more of them than that they move from place to place as they systematically denude the landscape of vegetation. Furthermore, they are usually stuffed full of bulky food to digest, which tends to produce inertia. Gigantopithecus probably followed this pattern. [Ciochon 1991]
The flexibility retained in their diet may be the result of recent specialization:
Chinese paleontologists are now speculating that the specialized bamboo diet of the living giant panda evolved rather recently in panda evolution, perhaps at the end of the Pliocene. [Ciochon 1990]
Bamboo belongs to the Gramineae family and consists of approximately 1,200 species of grass. It is long-lived, an evergreen woody grass, and varies greatly in size [Ciochon 1990]. Bamboo may have been part of the diet of Gigantopithecus [Ciochon 1990]. Bamboo has some unusual characteristics that can impact an animals ecology such as periodic die-offs. The last die-off during the 1970s was particularly hard on the giant panda as three major species of bamboo died-off simultaneously. During this period the giant panda turned to other food sources such as other plants and meat. Bamboo is found at altitudes of up to 11,500 feet in the Himalayas [Ciochon 1990].
The presence of fruit and grass phytoliths embedded in Gigantopithecus teeth suggests that the diet of Gigantopithecus may not have been as specialized as the giant panda's. We know that the giant panda, even with its specialized diet, will substitute alternate foods when bamboo is not available. Thus, since the giant panda has not yet become extinct as a result of the periodic die-off of bamboo, this probably did not play a significant role in the extinction of Gigantopithecus. Today anthropologists assert:
Gigantopithecus is the only ape known to have become extinct during the Pleistocene epoch. [Ciochon 1990]
Identifying the forces that drove Gigantopithecus to extinction can remove the attribution of Gigantopithecus as the fossil record of an uncataloged animal, thereby reducing the plausibility of an uncataloged animal.
Ecology
Examining the tenuous supposition that a descendant of Gigantopithecus is the source of the Bigfoot phenomenon requires the temporary suspension of disbelief.
This process is valuable in establishing the plausibility or implausibility of this supposition. The anecdotal reports are facts, as they tangibly exist — the supposition in question is whether the reports originate from the manifestation of an uncataloged animal, or if they should be attributed to a sociological phenomenon.
Habitat
The following information is extracted by subjective examination of the TBRP database of anecdotal observations and other anecdotal sources.
* Observations by westerners in the Rocky Mountains are principally from the 1800s with some 1700s observations.
* Observations by westerners in the Cascades are principally from the 1900s with some 1800s observations.
* Observations from the Himalayas and Asia are from three regions: Tibet, China and Russia.
Anecdotal observations similar to those of the Pacific Northwest also come from Canada, Alaska, Russia and China, all of which have a similar montane environment.
There is evidence that the giant panda, a contemporary of Gigantopithecus, migrated to the montane. Most anecdotal observations are in the higher and cooler montane environment such as the Cascades and Himalayas. One TBRP anecdotal report is from a glacier in the Cascades at approximately 9,000 feet.
It is estimated that the range of the giant panda has been cut in half during the last 140 years due to human encroachment. Two-hundred years ago, anecdotal observations originated from both the Pacific Northwest and the Rocky Mountains, which correlates with the position of the land gap between the North American ice sheets during the last ice age. Today, anecdotal observations originate principally from a narrow band in the Northwest, possibly the result of North American human encroachment.
Assuming an uncataloged animal exists, one reason why a type specimen has not yet been obtained may be the wide area of cover provided by large tracts of forested land in the Pacific Northwest. The US Forest Service, a division of the US Department of Agriculture, manages 141 million acres of national forest, 41 million of which are in Oregon, Washington and Idaho and account for more than 27% of the land mass of these states. Wilderness areas are contained within the national forests and their use is restricted: a use permit is required, there are no roads, there are no permanent man-made structures, all mechanized devices are prohibited. Five-percent of the combined land mass of Oregon, Washington and Idaho has this designation, which is an area larger than Massachusetts. There are also hundreds of thousands of acres of fully restricted land, such as watersheds, that no one may enter. The terrain in these areas is as rugged and inaccessible as areas in the Amazon and Himalayas. The population density of an animal (number per square kilometer) has been found to be allometrically related to body length (meters) [MacMahon 1983]:
Equation 9 (Eq. 9)
An alternative way of stating this is that population density is allometrically related to the amount of food consumed by an animal [MacMahon 1983]:
Equation 10 (Eq. 10)
The mean body length (stature) of the US office workers is 1.68 meters [Weimer 1993]. The Patterson-Gimlin film subject body length is approximately 2.2 meters. Sighting reports have estimated male stature as 2.7+ meters which when averaged yields an estimated mean of 2.45 meters, assuming an equal distribution of males and females in the population.
Diet
While unusual, anecdotal observations sometimes includes foraging information. TBRP reports include:
* Observation of foraging on cattails.
* Observation of foraging on bear grass, an evergreen grass that grows in clumps.
* Observation of stream fishing for steelhead (trout).
Ciochon believes Gigantopithecus may have eaten bamboo, which is found in the Himalayas up to 11,500 feet. The Himalayas are one source of contemporary anecdotal observations.
Bamboo and the Moraceae family, which may have been part of the diet of Gigantopithecus, are not native to the Pacific Northwest [USFS 1996]. Moraceae, commonly known as Mulberry, belongs to the Urticales. Also in the Urticale order and therefore closely related to Mulberry are: Elm (Ulmaceae), Hemp (Cannabaceae), and Nettle (Urticaceae). Nettle is native to the Pacific Northwest. While bamboo is not native, other grasses such as cattails and bear grass might substitute. There is also an abundance of fruits in the Cascades which might substitute for the Moraceae family.
Native forbs identified in early-seral vegetation plots on the Mt. Hood National Forest and Gifford Pinchot that are suitable for foraging include: Woodland beardtongue (Nothochelone nemorosa), Sitka burnet (Sanguisorba stichensis), Lupine (Lupinus latifolius), Cascade penstemon (Penstemon serrulatus), Bear grass (Xerophyllum tenax), Skunk cabbage (Lysichitum americanum), Cardwell's penstemon (Penstemon cardwellii), Great betony (Stachys cooleyae) and Sticky chickweed (Cerastium viscosum) [USFS 1996].
Kleiber's law expresses the relationship between mammalian body weight and energy requirements by an allometric relationship:
Equation 11 (Eq. 11)
where:
BMR is the basal metabolic rate,
k is a constant, and
WB is body weight [Jones 1992].
An exponent of 0.75 is appropriate for inter-species comparison, whereas 0.67 is appropriate for intra-species comparison.
Substituting the estimated body weight of the Patterson-Gimlin film subject of 887 kilograms, the BMR is 705 Watts. For comparison for the average world human weight of 57 kilograms the BMR is approximately 90 Watts.
Behavior
TBRP anecdotal reports of daytime sightings are at a maximum during the Pacific Northwest hunting season. There are many similar reports which enable the extraction of some common characteristics. A typical report is from hunters who, upon entering an area, or discharging their weapon, claims to see a Bigfoot stand up near them and walk away. Presumably, the noise of the approaching hunter, or from the weapon discharge, disturbs the subject. The giant panda spends approximately 40% of the day on it's back resting and eating. By ecological parallelism, resting and eating may be the source of the observed behavior in the anecdotal reports. Also, there are no TBRP anecdotal reports of a subject running. Also by ecological analogy, this is consistent with giant panda behavior which has never been seen running.
There are also similarities among TBRP anecdotal reports of night time sightings. A typical report is from a driver of a vehicle who claims to see a Bigfoot while driving on a road. Sometimes the subject crosses in front of the vehicle, sometimes it stands at the side of the road, and occasionally it crosses behind the vehicle. Often there are multiple people in the vehicle, and in at least one instance the incident involved a bus driver and a bus full of students. Night time sightings have led to speculation that the subject is nocturnal, but this is unlikely because all known nocturnal primates, except the aye-aye of Madagascar (Daubentonia madagascariensis) weigh less than 1 kilogram. Further, Delson argues that it is possible to judge if a primate is nocturnal or diurnal based upon orbit size and there are no TBRP anecdotal reports of a large orbit size [Delson 1985]. Nocturnalism may have separately evolved, but there would need to be a reason for the adaptation. No such reason has been identified at this time.
TBRP daytime and nighttime anecdotal reports, Green's Sighting Data, European settler records and Native American mythology indicate the subject is bipedal. There is no information in the fossil record to either support or contradict this. If the subject is pongid, and not hominid, the bipedalism may be the result of convergent evolution. If true, the reason for such an adaptation is unknown.
Cryptozoology
Conventional wisdom is that there are no new animals to be discovered, however there is a history of uncataloged animals. Linneaus began the process of formally cataloging the species in 1758. Figure 31 shows the trend of increasing numbers of cataloged species over time (birds, mammals, amphibians and reptiles only) [Heuvelmans 1958].
The 1817 survey is by Cuvier, 1886 by Leunis and Ludwig, 1898 by Mobius, and the 1939 survey is by Arndt [Heuvelmans 1958]. The 1960 data is an extrapolation made by Heuvelmans in 1958.
Fig. 31: Total Number of Cataloged Species by Year
Figure 31: Total Number of Cataloged Species by Year
Examples of "undiscovered animals" include:
* The 1869 discovery by the west of the giant panda which could not be located again for another 50 years. The first living specimen was obtained in 1936.
* The 1912 discovery of a type specimen of the Komodo dragon was which was thought to be mythical.
* The 1939 discovery of a live specimen of the coelacanth which was believed to be extinct for 70 million years.
* At one time it was generally accepted that the Precambrian was void of fossils. Today, paleontologists recognize fossilized stromatolites from the Precambrian as the remains of cyanobacteria, the oldest of which are from 3,000 million years ago. Recently, living stromatolites were discovered in Shark's Bay in western Australia [Fortey 1991].
The most strongly related example of an undiscovered animal is the lowland gorilla which parallels the Bigfoot phenomenon (Gorilla gorilla gorilla, Gorilla gorilla graueri). As early as 470 BC colonists from Carthage, who traveled to the West African coast, reported encountering hair-covered, stone-throwing animals. Other accounts are from 1774 and 1846. In 1847 the discovery of the gorilla is attributed to Savage and an observation from 1856 reports that gorilla are "never running from man..." [Kogod 1993]. Some of these accounts convey fearsome creatures and sound more mythical than observed. In the case of Chaillu, the exaggerations were introduced by his publisher. The resulting skewed perceptions were accepted as fact.
Humankind projects onto animals its desires and fears and in the end observes mainly the fiction it has created. In the black countenance and tremendous strength of the gorilla it sees less an animal than a myth, a mysterious and monstrous image of itself. [Richardson 1989]
After the discovery of the lowland gorilla reports persisted of yet another fearsome creature and were discredited. Then in 1902, the mountain gorilla was discovered (Gorilla gorilla berengi). Capable of living at both high and low elevations, they are often found between 7,800 and 11,000 feet in the Virunga Volcanoes located at the border of Zaire, Uganda and Rwanda. An important parallel between the gorilla and the Bigfoot phenomenon is:
There is no fossil record of chimpanzees or gorillas at all. [Jones 1992]
These parallels exist between the gorilla and the Bigfoot phenomenon:
Table 10: Gorilla and "Bigfoot" Parallels Mountain Gorilla Bigfoot
Mythology Thought to be myth until discovery Thought to be a myth
Morphology Standard for comparison Features similar to gorilla, but larger
Fossil Record None Limited (Gigantopithecus) or none
Ecology: Behavior Never runs No reports of running
Object throwing Reports of object throwing
Diet Omnivorous Reports of omnivorous diet
Habitat Montane Reported montane environment
BHM
Encounters with big hairy monsters have been reported continually, for millennia, from around the world. Reports come from credible and uncredible individuals alike, and may be reported immediately to authorities or, due to fear of ridicule, may be kept quiet for extended periods. While one "BHM" has been captured, the gorilla, the remainder are relegated to mythology.
BHM myths are thought to have existed for as long as humans have had language. Pliny reports an encounter with a BHM in 0 AD. There is no reason for us to expect that a universal myth that transcends cultures and time should disappear from use in modern times.
Carl Jung suggested that phenomenon such as BHMs are encounters with archetypal images derived from the collective unconscious, which are believed to be understood by all humans at a primitive level [Guiley 1995]. Our need for the BHM archetypal image may arise from a primal fear of the loss of control — a fear of encountering a force so overwhelming that the greatest of warriors could not overcome. The existence of such a primal fear may be an evolved survival strategy.
Brenda Sutherland, a graduate student of Ciochon who investigated anthropomorphic legends, addressed in her research the length of time myths survive. In some cases, tribal memory may extend tens-of-thousands of years. The bunyip, a mythical creature of the Aboriginal dreamtime may be a thirty-thousand year old tribal memory of the fossil Palorchestes that became extinct toward the end of the last Ice Age. There are similar records from other cultures.
Conclusion
The implausibility of an uncataloged animal may be demonstrated by the absence of a continuous record, since evolution theory generates an expectation of a continuous record of an organisms existence. The implausibility of an uncataloged animal may also be demonstrated through the implausibility of one or more dimensions of its ecology.
G. blacki is the only fossil record that approximates the size of anecdotal reports of Bigfoot. Four mandibles and approximately one-thousand teeth comprise the fossil record of Gigantopithecus, the youngest of which is from 400,000 to 200,000 years ago. The lack of G. blacki fossils in the intervening period, is partly why anthropologists believe G. blacki is extinct. There is however a 5 million year gap in the fossil record of Gigantopithecus and it seems presumptive to the author to interpret a 400,000 to 200,000 year gap in the fossil record as proof of the extinction of G. blacki.
Bamboo die-offs have been postulated as contributing to the extinction of G. blacki — this seems implausible given that its diet was probably more varied than the giant panda which survives to this day and which shared the ecology of Gigantopithecus.
There was a suitable environment for Gigantopithecus to migrate across the Bering Strait Land Bridge, and as evidenced by at least twenty-three other land mammals, cooperation and social structure were not required to make this crossing. Approximately 18,000 years ago a corridor of montane environment existed from the Bering Strait Land Bridge, between the Laurentian Ice Sheet and the western mountain ice, providing a montane conduit to the Rocky Mountains and Cascades.
G. blacki may be the fossil record of the Bigfoot phenomenon. It is also possible that like the gorilla, Bigfoot does not have a fossil record, as anecdotal reports come from the montane environment which does not support fossil formation. By itself, the lack of a fossil record is insufficient to demonstrate implausibility because of the example of a lack of a fossil record for the gorilla. There is no evidence that contradicts the possibility that G. blacki is the fossil record of the Bigfoot phenomenon. If it is extinct, "Gigantopithecus is the only ape known to have become extinct during the Pleistocene epoch." [Ciochon 1990]
Analysis of Native American stories and the GNIS search both show a bias toward the Pacific Northwest, paralleling Green's sighting data. The Native American stories may show a differentiation between a basal myth rate derived from stories of dwarfs, versus stories of big hairy monsters, indicating a higher frequency of Native American BHM myths than expected in the Pacific Northwest. Highpine demonstrated differences in Native American BHM myths, with Pacific Northwest myths sounding "observed" and the remainder more "mythical." Both the Native American stories and the GNIS search results could represent a regional memory or expectation of the phenomenon.
The analysis of observational data suggests it is the result of the superposition of multiple phenomenon. The analysis of Green's data suggests there may be willful manufacture by those reporting sightings. However, Green's data also shows two clusters, one in the Pacific Northwest which is differentiated from the remainder of the country. Willful manufacture is expressed as a basal rate of the population count and also accounts for some of the Pacific Northwest anecdotal reports.
Comparative morphology places the Patterson-Gimlin film subject between a mountain gorilla and a human. Analysis of the knee delta suggests locomotion differentiated from that of a human, although it is yet to be demonstrated that a human could not replicate the locomotion employed. Subsequent analysis of the knee range-of-motion data may be able to exclude the human archetype as a derivation for the film subject. As a single dimension, the Patterson-Gimlin film could be dismissed as a fake. However, forensic imaging analysis has been unable to relate the subject to the human archetype.
Perceptual failure accounts for a portion of the Bigfoot phenomenon, as there are documented instances where an individual identifies a tree trunk as a Bigfoot, or footprints as the result of double registration in bear prints, though there are only a small number of such occurrences in the TBRP database. There is probably a sociological basis for increased perceptual failure in the Pacific Northwest based upon the regional expectation of the phenomenon. The TBRP database also has instances of willful manufacture. However, there is no evidence that it is widespread.
Proof of the source of the Bigfoot phenomenon that is acceptable to the scientific community is the objective of this research. This may come from social science or psychological research into manufacturing and perceptual failure. In the event the phenomenon originates from an uncataloged animal, it is unfortunate, but nonetheless true, that anthropologists will demand a type specimen. By definition, the taxonomy of an uncataloged animal is unknown, which raises complex ethical and moral questions. To date, no type specimen of Bigfoot has been discovered, perhaps because it does not exist, but possibly because of the millions of acres of habitat and the natural disposal system in the montane environment — carcasses of known animals, such as bear, are rarely found. The continuity argument was unable to contradict the null hypothesis as there is at least one plausible model of continuity:
Fig. 32: Plausible Continuity Model
Figure 32: Plausible Continuity Model
It seems reasonable to attribute the Bigfoot phenomenon to anthropomorphic legend — perhaps Jungian in origin, derived from the collective unconscious, or perhaps a tribal memory of a now extinct animal. The aborigines do not, however, claim to have recently seen a bunyip whereas judges, sheriffs, police officers, and forest service employees claim to have recently seen a Bigfoot. In the opinion of the author, some of these individuals are credible, and are therefore less likely to fabricate a story or perpetrate a hoax. These people may have been a victim of a wide-spread hoax, or there may be wide-spread failure of perceptual mechanisms, but no evidence of this has been identified.
Willful manufacture, perceptual failure and sociological factors such as regional expectations are contributing sources to the Bigfoot phenomenon. Regional differences in anecdotal report frequencies and content differentiate the Pacific Northwest phenomenon from a North American basal BHM phenomenon. The origin of the Pacific Northwest phenomenon is as yet unidentified and may be of unknown social origin or perceptual failure, or it may be plausible that there is an uncataloged bipedal animal in the Pacific Northwest.
There is no hard evidence that proves the existence of Bigfoot, however, the quantity and distribution of anecdotal reports in the Cascades can not be readily discounted. Particularly compelling is the simultaneous presence of multiple dimensions of circumstantial evidence:
* plausible continuity,
* plausible ecology,
* Green's data,
* TBRP's data,
* GNIS data,
* Native American mythology,
* historical and contemporary anecdotal accounts, and
* the Patterson-Gimlin film,
which together, while coincidence has not yet been ruled out, suggest the presence of an underlying phenomenon. The lack of hard evidence supports the conclusion of anthropologists that Bigfoot does not exist and that Gigantopithecus is extinct. However, in the opinion of this author, the compelling circumstantial evidence warrants the dedication of additional resources to resolve the origin of the Bigfoot phenomenon.
Acknowledgments
I am indebted to many people who helped make this paper a reality. First and foremost, the staff of The Bigfoot Research Project: Peter Byrne, Tod Deery and Deborah Wolman, who on more than one occasion got me into trouble in the field, but more importantly, got me out alive. I would like to thank Tod and Deborah for their assistance preparing the TBRP data for this research, and Tod in particular for his tireless review of, and insightful comments on the early drafts of this paper.
My thanks to Pat Patterson1 for entrusting us with the Patterson-Gimlin Film, to Oxberry ATI for the fine digitization job they did, and to René Dahinden2 for granting us the right to incorporate the still images of the Patterson-Gimlin Film in this paper.
My sincere thanks to Henry Franzoni for his insights about Native American culture, and his willingness to contribute his Native American mythology and GNIS research to this paper.
Grover Krantz, whom I have never met, is the pioneer of the scientific investigation of this phenomenon and I thank him for laying much of the ground work upon which this research is based. John Green and John Napier have both performed work from which this paper has benefited. Robert Pyle observed "...the phenomenon of Bigfoot exists," which is the foundation for the framework upon which this paper is based. Dr. Richard Howe contributed many ideas to this paper including assisting with the formulation of the null hypothesis.
I would like to thank Dr. Robert Rines for his insatiable curiosity and his capacity to make the unknown, known. He has shown unswerving determination to find an answer to the mystery of this phenomenon.
Many people contributed to the content of this paper. I would also like to thank Henry Franzoni, Dr. William Saxe Wihr, Dr. Martin Lees, Melinda Scheuneman, Dr. Franklin Glickman, Peter Byrne, Dr. Robert Pyle, Dr. Howe for their assistance in editing this paper. Several people over several years assisted with the preparation of materials and processing of image data including: Dan Putnam, Stephanie Alsberg, James Deckert, James Barlow and David Pointer. My thanks to Henner Fahrenbach for his assistance with primatology research. My thanks to Chris Murphy for several interesting and useful conversations. My thanks also to Pasquale and Jacquie Barone and the entire staff of the Hood River Hotel who are warm and gracious hosts and always made me feel at home during my many visits to Hood River, Oregon.
My special thanks to Melinda Scheuneman who, for several weeks, was my eyes and ears performing research in the library, and who somehow managed at the same time to keep the office running in my absence.
Vitae: Jeff Glickman received his Bachelor of Science in Computer Science from the University of Illinois at Urbana-Champaign in 1982. While there he the was Assistant Director of the Information Engineering Laboratory and the Director of Software Development for the Computer Research Laboratory. While at the University he worked extensively with imaging and optical systems.
Mr. Glickman has developed proprietary advanced image processing techniques and has been called upon to apply them to solve criminal cases for law enforcement agencies including the NYPD, US Attorney's Office and the FBI, as well as litigations and scientific investigations. Mr. Glickman is a Board Certified Forensic Examiner.
Correspondence may be directed to Mr. Glickman at Photek, 209 Oak Street, Suite 202, Hood River, Oregon, 97031.
1. Mrs. Patterson was compensated for granting this research project access to the film.
2. René Dahinden was compensated for granting this research project access to his still images.
Appendix A
Speculation
A tenuous string of suppositions derived from the anecdotal observations and from the ecological analogy with the giant panda and the orangutan might narrow a search for those wishing to try. For an uncataloged animal to be plausible, each dimension of its ecology must also be plausible.
The following are based upon a combination of fossil evidence and anecdotal observations and is based upon current knowledge and information. The subject, if it exists, may be (or have):
* Pongid
* Diurnal
* Bipedal
* Montane habitat
* Omnivorous diet consisting of grass, fruit, fish
* Slow, probably never running
* Shy, walking away upon encountering a human
* Spends most of the day resting and eating
* Seven to nine feet in stature
* Possibly 1,900 lb.+ mass (estimated)
The current interpretation of the fossil evidence suggests Gigantopithecus was pongid, which, if it is the fossil record of the Bigfoot phenomenon, suggests that Bigfoot is pongid. Morphological comparisons derived from the Patterson-Gimlin film are in agreement with this and suggest that Bigfoot is related to the mountain gorilla and is therefore pongid.
Appendix B
Recommendations for future study
The following are recommendations for research into the Bigfoot phenomenon:
* Independent verification of results
* Study of human attempting gait observed in Patterson-Gimlin film
* Extraction of jaw and tooth size from Patterson-Gimlin film for comparison with Gigantopithecus fossils
* Test anecdotal observations with theory of testimony
* Studies to test for sociological origin
* Studies to test for perceptual failure
* Statistical analysis of sighting data integrated with ecological data (flora, etc.)
* Integration of all available observational data into a single database (Green, TBRP, etc.)
* Deployment of advanced technology for automated wildlife surveys
Appendix C
Names of big hairy monsters
Table 11: World-Wide Names (Excluding North America) Name Country
Abominable Snowman Tibet
Agogue East Africa
Alma Mongolia
Almas Mongolia
Almasty Russia
Cigouve Haiti
Duendi Columbia
Dzestarnacks China
Homo ferus Sweden
Homo nocturnus Sweden
Homo sylvestris Sweden
Homo troglodytes Sweden
Metah Kangmi Nepal
Orang-Dalam Malaysia
Orang-Pendek Borneo
Quidili Russia
Sedapa Borneo
Shookpa Nepal
The Big Grey Man Scotland
Yeh Ren China
Yeti Nepal, Tibet
Yowie Australia
Table 12: Native American Names (North America) Name Tribe Description
A-hoo-la-huk Yup'ik Apotamkin Maliseet-Passama-quoddy A bogey monster with long hair and huge teeth. Fear of him keeps small children from straying onto thin, newly frozen ice in the winter and unguarded beaches in the summer
Asin Alsea A fearful monster-girl who lives in the woods and carries people off, especially unattended children
At'at'ahila Chinookan
Atahsaia Zuni
Big Figure Kwakwaka'wakw
Big Hairy Man Hopi
Boqs Bella Coola
Bukwas Kwakwaka'wakw Wild Man of the Woods
Bushmen Hare
Chahnameed Pequot The great eater, the glutton. A giant who lives alone on an island and lusts after a beautiful women walking along the beach of the mainland
Chiha-tanka Dakota Sioux big elder brother
Chiye-tanka Lakota Sioux big elder brother
Dehotgohsgayeh Onondaga, Iroquois This giant lives in the south along the margin of the earth in total darkness. One side of his body is red, the other black
Desini Chilcotin Strangers, rarely seen who steal women. Desini are found in the vicinity of camping places
Dzonoqua, Tsonaqua Kwakwaka'wakw Wild Woman of the Woods, An ugly giantess who steals children
Free-man various modern Native American tribes
Fsti capcaki Seminole A giant covered with gray hair who smells like a stagnant muddy pond
Gagixit Haida Wild men. Men who were made wild by the Land of the Otter people
Geldegwests Coos Benign giants who live near streams and eat fish
Get'qun Lake Iliamna Athabas-can
Gilyuk Nelchina Plateau The big man with the little hat
Goo-tee-khl Tinglit, Chilkat
Hairy Man Tanaina A large, harmless, hair covered creature who lives in mountain villages. The eyes of this grayish, two-legged being have no pupils. He is helpful to a human unless injured
Indacinga Ponca Beings with great physical strength who live the forests and hoot like owls. Mothers use the threat of being caught by Indacinga to influence the behavior of their children
nugpasugssuk Eskimo A giant who catches fish and seals with his bare hands
Iya Lakota The malevolent giant created by Inyan after Skan and Maka
Kala'litabiqw Skagit Valley
Kashehotapolo Choctaw A beast-man with a shriveled head
Katyutayuuq Eskimo A female monster with no body and whose large head is attached directly to her feet
Kauget Coast Salish
Kiwahkw Maliseet-Passama-quoddy Cannibal ice giants. The corpse of one witch killed by another
Kushtaka Tlingit
Loo-poo-oi'yes Miwuk
Miitiipi Kawaiisu
Nahgane Slavey Bush giants who steal careless young children
Nant'ina Dena'ina Athabascan
Nantiinaq Kenai Penisula Native Americans
Neginla-eh Alutiiq, Yukon Wood Man
Nulayuuiniq Eskimo A newborn female who suddenly grows to be a giant
Olayome Native Americans near Clear Lake California
Omah Yurok
Ot-ne-yar-heh, Stonish Giants Iroquois
Qah-lin-me Yakama, Klickitat
Qaxdascidi Tanaina A malevolent being known by the mysterious noises it makes
Qui-yihahs Yakama, Klickitat The five brothers
Rugaru Ojibway
Saskehavas Coast Salish
Sasquatch
Seahtik, Selatik, Seeaht-kch Clallam
Seat-ka Yakama
Seatco Puyallup, Nisqually A malevolent, larger-than-human figure known for his stealth and quickness
See'atco Coast Salish One who runs and hides
Shadow Indians Yakama
Skookum, Scoocum Chinook Evil God of the Woods
Sne-nah Okanogan Owl Women
So'yoko, Si'Yoko Hopi, Lakota Sioux
Spirit, Spirit of the Woods many Native American tribes
Ste-ye-hah, Ste-ye-mah Yakama Spirit hidden under the cover of the woods
Steta'l Puyallup, Nisqually Spirit Spear
Stick-Shower, Stick Indians Yakama, Klickitat, Puyallup, Puget-Sound, Colville
T'oylona Taos person big
Tah-tah-kle'-ah Yakama, Shasta Owl Woman Monster
Tammatuyuq Eskimo An infant-killing monster who lives in the time of the first people
The Big Man Oglala Lakota Sioux
The Hairy Man Alaskan Athabascan
The Stone Giants, Stone Coats, Ge-no-sqwa Seneca
Tsavoojok Paviotso An old giant who challenges husbands to fight one another so he can steal their wives
Tsiatko Puyallup, Nisqually
Tso-apittse Shoshone
Tuurnngaq Eskimo Ancient giant humans who live in solid rock houses. They kill people and cause hunters to disappear
Urayuli Southwest Alaskan Eskimo
Wakandagi Omaha, Ponca Long-bodied, horned monsters
Wetiko Cree
Winalagilis Kwakiutl A giant who travels the world in a canoe he never leaves
Windago Eastern Athabascan
Witiko non-human giant, filthy, mean appearance
Xi'lgo Nahalem Tillamook Wild Woman
Yahyahaas Modoc
Yi'dyi'tay Nehalem Tillamook Wild Man
Wild Man, Wild Man of the Woods Arkansas
Yeahoh Kentucky
Table 13: European Settler Names (North American) Name Location/Description
Bigfoot California
Boogers
Cape Apes Oregon Coastline
Devil Demon, Mountain Devil Western United States
Diablo Western United States (Spanish)
Grey Man North Carolina, South Carolina
Mo-Mo Kansas, Missouri
Old Sheff Kansas, Missouri
Skunk Apes Washington, Florida
Splintercat Oregon
Swamp Monsters
The Old Man of the Crater Washington
The Snow Walker Roosevelt, Theodore (story)
Wampus, Wampus Cat North Carolina, Oregon
Wild Man, Wild Man of the Woods Arkansas
Yeahoh Kentucky
Appendix D
Human segment dimensions
Table 14: Human Segment Dimensions Segment Segment Length as Percentage of Height
Head Length 0.130H
Shoulder-to-Shoulder Width 0.259H
Upper Arm Length 0.186H
Forearm Length 0.146H
Hand Length 0.108H
Chest Width, Frontal (not circumference) 0.174H
Hip Width 0.191H
Foot Width 0.055H
Foot Length 0.152H
Ankle Height (from ground) 0.039H
Knee Height 0.285H
Hip Height 0.530H
Chest Height 0.720H
Fingertip Height 0.377H
Wrist Height 0.485H
Elbow Height 0.630H
Shoulder Height 0.818H
Chin Height 0.870H
Bibliography
* Barnes-Svarney, Patricia Ed., The New York Public Library Science Desk Reference, Macmillan, New York, New York, 1995.
* Bloem, Doug, Telephone call, Dalles Watershed, Oregon, 1996.
* Bronstein, Daniel A., Law For the Expert Witness, Lewis Publishers, Boca Raton, Florida, 1993.
* Burenhult, Goran, Ed., The First Humans: Human Origins and History to 10,000 B.C., HarperCollins Publishers, New York, New York, 1993.
* Bryant, Page, Native American Mythology, Aquarian HarperCollins Publishers, London, England, 1991.
* Byrne, Peter, Personal Conversations, Telephone Calls and Correspondence.
* Byrne, Peter, The Search for Bigfoot: Monster, Myth or Man?, Acropolis Books, Ltd., Washington, D.C. 1975.
* Byrne, Peter, Hodgson, Michael, et. al., Research Photography of the Bluff Creek Site, 1972.
* Calder, Nigel, The Weather Machine, The Viking Press, New York, New York, 1974.
* Carr, Joseph J., The Art of Science, Hightext Publications, Inc., San Diego, California, 1992.
* Cavalli-Sforza, L. Luca, Menozzi, Paolo and Piazza, Alberto, The History and Geography of Human Genes, Princeton University Press, Princeton, New Jersey, 1994.
* Chellappa, Rama, Ed., Digital Image Processing, IEEE Computer Society Press, Los Alamitos, California, 1992.
* Chivers, David J., Wood, Bernard A. and Bilsborough Alan, Eds., Food Acquisition and Processing in Primates, Plenum Press, New York, New York, 1984.
* Chui, Charles K., Montefusco, Laura and Puccio, Luigia, Editors, Wavelets Theory, Algorithms and Applications,Academic Press, San Diego, California, 1994.
* Ciochon, R. L., Hominoid Cladistics and the Ancestry of Modern Apes and Humans: A Summary Statement, New Interpretations of Ape and Human Ancestry, Plenum Press, pp. 783-843, 1983.
* Ciochon, Russel, Telephone Call, 1995.
* Ciochon, Russell, Olsen, John and James, Jamie, Other Origins, Bantam Books, New York, New York, 1990.
* Ciochon, Russell L., The Ape That Was, Natural History, pp 54-62, Nov. 1991.
* Ciochon, Russell and Piperno, Dolores, Scratching the surface of evolution, New Scientist, pp 47-49, Nov. 1990.
* Ciochon, Russell, Piperno, Dolores R. and Thompson, Robert G., Opal phytoliths found on the teeth of the extinct ape Gigantopithecus blacki: Implications for paleodietary studies, Proc. Natl. Acad. Sci., Vol. 87, pp 8120-8124, 1990.
* Clutton-Brock, T. H., Ed., Primate Ecology: Studies of feeding and ranging behaviour in lemurs, monkeys and apes, Academic Press, London, England, 1977.
* Cook, Thomas D. and Campbell, Donald T., Quasi-Experimentation, Design & Analysis Issues for Field Settings, Houghton Mifflin Company, Boston, Massachussetts, 1979.
* De Boer, Leobert E.M., Ed., The Orang Utan: Its Biology and Conservation, Dr. W. Junk Publishers, The Hague, Netherlands, 1982.
* Delson, Eric, Ed., Ancestors: The Hard Evidence, Alan R. Liss, Inc., New York, New York, 1985.
* Embree, Paul M., C Algorithms for Real-Time DSP, Prentice-Hall PTR, Upper Saddle River, New Jersey, 1995.
* Embree, Paul M. and Kimble, Bruce, C Language Algorithms for Digital Signal Processing, Prentice Hall, Englewood Cliffs, New Jersey, 1991.
* Evans, Charlotte Ed., The Kingfisher Illustrated History of the World, Kingfisher Books, New York, New York, 1992.
* Famighetti, Robert, Ed., The World Almanac and Book of Facts 1996, Funk & Wagnalls Corp., Mahwah, New Jersey, 1995.
* Farhenbach, Henner, Personal Correspondence, Conversations and Telephone Calls, 1995-1996.
* Fortey, Richard, Fossils: The Key to the Past, Harvard University Press, Cambridge, Massachusetts, 1991.
* Franzoni, Henry, Legends Beyond Psychology, Revision 4.4, unpublished, 1996.
* Franzoni, Henry, GNIS Search, unpublished, 1996.
* Franzoni, Henry, Personal Correspondence, 1995-1996.
* Galdikas, Birute M. F., Reflections of Eden: My years with the Orangutans of Borneo, Little Brown and Company, Boston, Massachusetts, 1995.
* Gill, Sam D. and Sullivan, Irene F., Dictionary of Native American Mythology, Oxford University Press, New York, New York, 1992.
* Glickman, Jeff, NASI: Patterson-Gimlin Film Research Results, Internal Report of the North American Science Institute, 1997.
* Godwin, Sara, Gorillas, Friedman/Fairfax Publishers, New York, New York, 1994.
* Gonzalez, Rafael C, and Wintz, Paul, Digital Image Processing, 2nd Ed., Addison-Wesley Publishing Company, Reading, Massachusetts, 1987.
* Green, John, Sasquatch, The Apes Among Us, Hancock House Publishers, Inc., Blaine, Washington, 1981.
* Green, John, Telephone Calls, 1996.
* Guiley, Rosemary Ellen, Atlas of the Mysterious in North America, Facts on File, Inc., New York, New York, 1995.
* Hamburg, David A. and McCown, Elizabeth R., Eds., The Great Apes, The Benjamin/Cummings Publishing Company, Menlo Park, California, 1979.
* Harrison, Barbara, Orang-utan, Doubleday & Company, Inc., Garden City, New York, 1963.
* Heuvelmans, Bernard, On the Track of Unknown Animals, Richard Clay and Company, Ltd., Bungay, Suffolk, Great Britain, 1955.
* Hodgson, Michael, Telephone Call with Peter Byrne, 1995.
* Hudson, Ralph, The Engineer's Manual, 2nd. Ed., John Wiley and Sons, New York, 1917.
* James, Jamie, Stalking the Giant Ape, Discover, pp. 43-50, Feb. 1989.
* Johnson, Jr., A. E., Skeletal Estimates of Gigantopithecus Based on Gorilla Analogy, Journal of Human Evolution, Vol. 8, pp. 585-587, 1979.
* Jones, Steve, Martin, Robert and Pilbeam, David Eds., The Cambridge Encyclopedia of Human Evolution, Cambridge University Press, 1992.
* Kodak Archives, Telephone Call, Rochester, New York, 1995.
* Kogod, Charles Ed., The Great Apes, National Geographic Society, Washington, D.C., 1993.
* Krantz, Grover S., Anatomy of the Sasquatch Foot, Northwest Anthropological Notes, Vol. 6, No. 1, pp. 91-104.
* Krantz, Grover S., Big Footprints, Johnson Books, Boulder, Colorado, 1992.
* Lankford, Nancy, Telephone call, Bull Run Watershed, Oregon, 1996.
* Leakey, Richard, The Origin of Humankind, BasicBooks HaperCollins Publishers, New York, New York, 1994.
* Lindley, Craig A., Practical Image Processing in C, John Wiley & Sons, Inc., New York, New York, 1991.
* Maple, Terry L., Orang-utan Behavior, Van Nostrand Reinhold Company, New York, New York, 1980.
* Martin, R. D., Primate Origins and Evolution: A phylogenetic reconstruction, Chapman and Hall, Ltd., London, England, 1990.
* Matson, Jack V., Effective Expert Witnessing 2nd Ed., Lewis Publishers, Boca Raton, Florida, 1994.
* Mattison, Phillip E., Practical Digital Video With Programming Examples in C, John Wiley & Sons, Inc., New York, New York, 1994.
* McMahon, Thomas A. and Bonner, John Tyler, On Size and Life, Scientific American Books, Inc., 1983.
* Morris, Ramona and Morris, Desmond, The Giant Panda, Kogan Page, Ltd., London, England, 1981.
* Napier, J. R. and Napier, P. H., A Handbook of Living Primates: Morphology, Ecology and Behavior of Non-human Primates, Academic Press, London, England, 1967.
* Napier, John, Bigfoot: The Yeti and Sasquatch in Myth and Reality, The Trinity Press, London, England, 1972.
* Neufeldt, Victoria Ed., Webster's New World College Dictionary 3rd Ed., Macmillan, New York, New York, 1996.
* New York Times (1994) "Tests Show Giant Deer is a New Species", AP Wire Service, April 26, 1994.
* Nichols, Michael and Schaller, George B., Gorilla, Apeture, New York, New York, 1989.
* Nigg, Benno M. and Herzog, Walter, Eds., Biomechanics of the Musculo-skeletal System, John Wiley & Sons, New York.
* Patterson, Roger and Gimlin, Bob, "1967 Patterson-Gimlin Film", 16mm motion picture, Bluff Creek, California, 1967.
* Parker, J. R., Practical Computer Vision Using C, John Wiley & Sons, Inc., New York, New York, 1994.
* Pavlidis, Theo, Algorithms for Graphics and Image Processing, Computer Science Press, Rockville, Maryland, 1982.
* Phillips, Dwayne, Image Processing, R&D Publications, Inc., Lawrence Kansas, 1994.
* Piattelli-Palmarini, Massimo, Inevitable Illusions, John Wiley & Sons, New York, New York, 1994.
* Pilbeam, David, Gigantopithecus and the Origins of Hominidae, Nature, Vol. 225, pp. 516-519, 1970.
* Pitas, Ioannis, Digital Image Processing Algorithms, Prentice Hall, New York, New York, 1993.
* Pratt, William K., Digital Image Processing, 2nd Ed., John Wiley & Sons, Inc., New York, New York, 1991.
* Pyle, Robert Michael, Where Bigfoot Walks, Houghton Mifflin Company, New York, New York, 1995.
* Redsicker, David R., The Practical Methodology of Forensic Photography, CRC Press, Boca Raton, Florida, 1994.
* Reid, Cristopher E. and Passin, Thomas B., Signal Processing in C, John Wiley & Sons, Inc., New York, New York, 1992.
* Richardson, Nan, Ed., Gorilla, Struggle for Survival in the Virungas, Aperture Foundation, Inc., New York, New York, 1989.
* Robbins, Louise E., Footprints, Charles C. Thomas Publisher, Springfield, Illinois, 1985.
* Rorabaugh, C. Britton, Digital Filter Designer's Handbook, TAB Books, Blue Ridge Summit, Pennsylvania, 1993.
* Russ, John C., The Image Processing Handbook, CRC Press, Inc., Boca Raton, Florida, 1992.
* Sapunov, Valentin B., A Mathematical Analysis of "Snowman" Eyewitness Reports, Cryptozoology, Vol. 7, pp 61-65, 1988.
* Scarre, Chris, Smithsonian Timelines of the Ancient World, Dorling Kindersley, London, England, 1993.
* Schaller, George B., The Mountain Gorilla: Ecology and Behavior, The University of Chicago Press, Chicago, Illinois, 1963.
* Schaller, George B., Jinchu, Hu, Wenshi, Pan and Jing, Zhu, The Giant Pandas of Wolong, The University of Chicago Press, Chicago, Illinois, 1985.
* Schwartz, Jeffrey H., The Red Ape: Orang-utans and Human Origins, Houghton Miffline Company, Boston, Massachusetts, 1987.
* Sheldon, William G., The Wilderness Home of the Giant Panda, University of Massachusetts Press, Amherst, Massachusetts, 1975.
* Sid-Ahmed, Maher A., Image Processing, McGraw-Hill, Inc., New York, New York, 1995.
* Simons, Elwyn L. and Ettel, Peter C., Gigantopithecus.
* Smuts, Barbara B. and Cheney, Dorothy L, et. al., Eds., Primate Societies, The University of Chicago Press, Chicago, Illinois, 1986.
* Soumekh, Mehrdad, Fourier Array Imaging, PTR Prentice Hall, Englewood Cliffs, New Jersey, 1994.
* Sussman, Robert W., Primate Ecology: Problem-oriented field studies, John Wiley & Sons, New York, New York, 1979.
* Sprague, Roderick and Krantz, Grover S., Eds., The Scientist Looks at Sasquatch II, The University Press of Idaho, Moscow, Idaho, 1979.
* Tattersall, Ian, The Human Odyssey: Four Million Years of Human Evolution, Prentice Hall General Reference, New York, New York, 1993.
* Turekian, Karl K., Ed., The Late Cenozoic Glacial Ages, Yale University Press, New Haven, Connecticut, 1971.
* Unknown, Sixes Wild Man Again, The Lane Country Leader, Cottage Grove, Oregon, Vol XV., No. 51, 1904.
* U.S. Department of the Interior, Geographic Names Information System (GNIS CD-ROM), 1994.
* Weimer, Jon, Handbook of Ergonomic and Human Factors Tables, PTR Prentice Hall, Englewood Cliffs, New Jersey, 1993.
* Winter, David A., Biomechanics and Motor Control of Human Movement 2nd Ed., John Wiley & Sons, New York, New York, 1990.
* YinYun, Zhang, Variability and Evolutionary Trends in Tooth Size of Gigantopithecus blacki, American Journal of Physical Anthropology, Vol. 59, pp 21-32, 1982.
Part 1 | Part 2 | Part 3
Download this paper (1.5MB, PDF format; right-click and choose 'Save As')
The film that Roger Patterson and Bob Gimlin made in 1967 is real. Bigfoot is a human-primate hybrid.
ReplyDeleteBigfoot was created by men that had sex with female gorillas and orangutans a long time ago.
http://www.youtube.com/watch?v=
QBcxquJ1J6w&feature=fvwrel